Вектор золотое сечение: D0 b7 d0 be d0 bb d0 be d1 82 d0 be d0 b5 d1 81 d0 b5 d1 87 d0 b5 d0 bd d0 b8 d0 b5 d0 b2 d0 b5 d0 ba d1 82 d0 be d1 80: стоковые векторные изображения, иллюстрации
22.11.1974 


 Разное
Разное
Золотое сечение Иллюстрация ai svg
Золотое сечение Иллюстрация ai svgключевые слова
- фон
- баланс
- сочинение
- концепция
- дизайн
- Фибоначчи
- формула
- геометрический
- геометрия
- золотой
- Золотое сечение
- гармония
- значок
- иллюстрация
- математика
- номер
- Пропорции
- соотношение
- раздел
- знак
- спираль
- квадрат
- Водоворот
- символ
- симметрия
- технологии
- шаблон
- Золотое сечение Иллюстрация
- ai
- svg
- Золотое
- сечение
- Иллюстрация
DMCA Contact Us
бесплатная загрузка ( ai svg, 1.92MB )Связанная векторная графика
- Вектор иллюстрации золотого сечения svg eps
- Вектор золотого сечения svg
- Вектор иллюстрации золотого сечения svg eps
- Файл эскиза золотого сечения sketch
- Файл эскиза золотого сечения sketch
- Золотое Сечение Фибоначчи Шаблон Логотипа Векторные Иллюстрации ai svg eps
- Вектор иллюстрации золотого сечения svg eps
- Золотое сечение шаблон логотипа векторные иллюстрации svg eps ai
 org/ImageObject»> Вектор золотого сечения svg
org/ImageObject»> Вектор золотого сечения svg- Файл эскиза золотого сечения sketch
- Геометрия золотого сечения svg ai
- Лавровый венок Золотой векторные иллюстрации ai
- Современные границы векторные иллюстрации в стиле Версаче с золотым старинным греческим ключом eps svg
- Золотое сечение Иллюстрация ai svg
 org/ImageObject»> Иллюстрация Золотой якорь ai eps
org/ImageObject»> Иллюстрация Золотой якорь ai eps- Формирование золотого сечения svg ai
- Золото с вектором градиента розового золота eps
- Современные границы векторные иллюстрации в стиле Версаче с золотым старинным греческим ключом svg eps
- Черно-золотая визитка eps
- Красивый золотой фон ai eps
- Современные границы векторные иллюстрации в стиле Версаче с золотым старинным греческим ключом eps svg
 org/ImageObject»> Золотой лавровый венок с золотыми лентами векторная иллюстрация ai
org/ImageObject»> Золотой лавровый венок с золотыми лентами векторная иллюстрация ai- Векторный набор золотых градиентов ai
- Золотая Корона eps ai
- Роскошный декоративный фон мандалы в золотом цвете eps
- Золотой значок и рамка eps ai
- Золотая лента узор векторной графики eps
- Румяна текстуры розового золота
 org/ImageObject»> Золотая звезда значок psd
org/ImageObject»> Золотая звезда значок psd- Золотой лев вектор eps
- Золотое украшение мандалы ai
- Коллекция золотых лент eps
- Набор золотых световых эффектов eps
- Золотые текстовые эффекты psd
- Золотые декоративные элементы eps
- Золотой открывающий баннер eps
- Золотая цветочная роскошная рамка eps
- Золотая старинная рамка с цветами eps
- VIP золотой логотип eps
- Роскошные золотые этикетки векторный материал eps
- Золотая бумага загнутые углы векторные иллюстрации eps
- Серо-золотая визитка ai eps
- Золотая металлическая текстура eps
- Золотой декоративный логотип eps
 org/ImageObject»> Элегантная золотая рамка-орнамент eps
org/ImageObject»> Элегантная золотая рамка-орнамент eps- 3d золотой текстовый эффект psd
- Contact Us
%d0%b7%d0%be%d0%bb%d0%be%d1%82%d0%be%d0%b5 %d1%81%d0%b5%d1%87%d0%b5%d0%bd%d0%b8%d0%b5 PNG, векторы, PSD и пнг для бесплатной загрузки
естественный цвет bb крем цвета
1200*1200
схема бд электронный компонент технологии принципиальная схема технологическая линия
2000*2000
green environmental protection pattern garbage can be recycled green clean
2000*2000
be careful to slip fall warning sign carefully
2500*2775
3d модель надувной подушки bb cream
2500*2500
три группы 3d реалистичное декоративное яйцо с золотым цветом на гнезде bd с золотым всплеском текстовый баннер
5000*5000
be careful to fall prohibit sign slip careful
2300*2600
black key that can be hung on the body car key key
2000*2000
black and white train icon daquan free download can be used separately can be used as decoration free of charge
2000*2000
bb крем ню макияжа постер Новый список преимущественный колос День святого
3240*4320
крем крем вв вв на воздушной подушке иллюстрация
2000*2000
flowering in summer flower buds flowers to be placed lotus
2000*2000
childrens day childrens day border childrens day cute border can be used for childrens day theme poster stickers
842*596
chinese wind distant mountain pine tree chinese style pine tree chinese style poster can be combined
3600*2475
две бутылки косметики жидкая основа белая бутылка крем bb
2000*2000
logo design can be used for beauty cosmetics logo fashion
1024*1369
элегантный серебряный золотой bb позже логотип значок символа
1200*1200
bb крем ню макияж косметика косметика
1200*1500
в первоначальном письме bd логотипа
1200*1200
beware of being down be careful pay attention to safety warning signs
2000*2559
цвет перо на воздушной подушке bb крем трехмерный элемент
1200*1200
облака комиксов
5042*5042
be careful of electric shock signs warning signs warnings
2000*2000
bd письмо логотип
1200*1200
в первоначальном письме bd шаблон векторный дизайн логотипа
1200*1200
номер 82 золотой шрифт
1200*1200
bd письмо 3d круг логотип
1200*1200
bb крем cc крем пудра Порошок торт фонд
2000*2000
flowering in spring flower buds flowers to be placed rhododendrons
2000*2000
2022 календарь bd с фоторамкой
2500*2500
глюк числа 87 вектор на прозрачном фоне
1200*1200
жидкая подушка крем bb
1200*1200
be careful warning signs warning signs be
2000*2000
be careful of potholes warning signs warning signs caution
2000*2000
big cock detailed layered can be used directly cock big cock chicken
2000*2000
black and white eco friendly pattern garbage can be recycled green clean
2000*2000
bb логотип письмо дизайн вектор простые и минималистские ключевые слова lan
1202*1202
prohibited use mobile phone illustration can not be used
2048*2048
простая инициализация bb b геометрическая линия сети и логотип цифровых данных
2276*2276
аэрозольный баллончик увлажняющий лосьон bb cream парфюм для рук
3072*4107
в первоначальном письме bd шаблон векторный дизайн логотипа
1200*1200
3d золотые числа 82 с галочкой на прозрачном фоне
1200*1200
3d золотые числа 87 с галочкой на прозрачном фоне
1200*1200
Креативное письмо bb дизайн логотипа черно белый вектор минималистский
1202*1202
Красивая розовая и безупречная воздушная подушка bb крем косметика постер розовый красивый розовый Нет времени На воздушной
3240*4320
на воздушной подушке на воздушной подушке bb крем консилер отрегулировать тон кожи
2000*2000
номер 82 3d рендеринг
2000*2000
номер 81 золотой шрифт
1200*1200
3d золотые числа 81 с галочкой на прозрачном фоне
1200*1200
flowering in spring flower buds flowers to be placed rose
2000*2000
Рассказ «Золотое сечение» — Вектор-успеха.
 рф
рф— Два билета первого класса, пожалуйста… Опять востребован в другой части света, да, вечные перелеты… помогаю даже архитекторам (их последнее время как собак нерезанных) … Ха-ха, представьте себе: зазнался на столько, что допустил подобное высказывание! Позор, да и только. А раньше был счастлив поработать с египетскими строителями (их в ту пору тоже немало было) …
— Что, простите? А, да-да, в строительстве пирамиды Хеопса. Чуть позже сотрудничал с греками: участвовал в создании Парфенона да увлекался по мелочи скульптурами, а они в свою очередь причисляли меня чуть ли не к главным создателям мира, что, к слову сказать, было мне на руку: мог балагурить и буянить, сколько душе угодно, а они меня все равно почестями баловали…Эх, хорошие были времена… С ними только эпоха Возрождения может сравниться: меня тогда называли “божественным”, представляете!
— Смеетесь? И мне смешно, до чего люди любят преувеличивать значение помощи. Подумаешь, приукрасил лицо Джоконды и некоторых других известных дам, но не “божественным” же сразу называть, ей-богу.
— Да, это чистая правда. С Шопеном и Бетховеным лично за руку здоровался, с вашим Чайковским состоял в самых дружеских отношениях…
— Перестаньте Вы ахать, дамочка, простейшие (раз уж высшая математика отказывается меня воспринимать) законы математики и Вас могли бы сделать культовым композитором, а если захотите, то и поэтом (да-да, Пушкин и Руставли тоже мои подопечные). Вы только попросите… Но не в этом веке, я слегка занят наукой. Может быть, когда возьму отпуск…
— Какой наукой? Самой современной!
— Да что там высшая математика? Не только она определяет вектор прогресса, уж поверьте мне! Знаете об открытии квазикристаллов? А о филлотаксисе? О законе структурной гармонии систем? О структуре ДНК? — это все малая часть моих заслуг в современном обществе!..
— Девушка, простите, вот и мой помощник Ч. Фибоначчи, без него не обходилось ни одно мое открытие, мы не можем пропустить самолет: люди из сферы бизнеса и торговли невероятно нервные. А этот цветок для Вас, на память, к слову, тоже мое творение. Возможно, во мне действительно есть что-то божественное… До свидания.
Возможно, во мне действительно есть что-то божественное… До свидания.
Только что перед агентом воздушных сообщений прошла идеальная пропорция, чья наименьшая часть так относится к наибольшей, как наибольшая ко всему. Как настолько простое явление может играть такую важную роль в процессе мироустройства? Где еще затаилось золотое сечение и почему не раскрывает всех своих загадок, что нужно сделать, чтобы обнаружить его?
В темном переулке раздается тихий звук шагов. Фонарь рисует фигуру человека на многоэтажках города. Его развивающийся плащ укрывает спящих жителей. Как всегда, он прошел незамеченным мимо сотен людей, слишком озабоченных повседневными делами. Человек, который хранит покой всего мира, заходит в свою одинокую квартиру, берет газету, пытающуюся разгадать тайну Золотого сечения, его тайну, и смеется:
— Просто оглядитесь. Я среди вас.
Векторы в развитии талантов и способностей у детей и молодёжи
29 сентября Юрий Биктуганов на базе Регионального модельного центра Свердловской области, структурного подразделения Дворца молодёжи, провёл расширенное заседание Министерства образования и молодёжной политики Свердловской области «Об организации работы по выявлению, поддержке и развитию способностей и талантов у детей и молодёжи Свердловской области».
В событии приняли участие руководители городских и региональных организаций, заинтересованных в подготовке проектов по вопросам развития способностей обучающихся, методического сопровождения педагогов, повышения доступности и качества услуг дополнительного образования, организации региональной системы выявления талантов.
Для всех участников заседания была проведена экскурсия по Региональному модельному центру. Созданный в 2018 году в рамках реализации одного из направлений реализации федерального проекта «Успех каждого ребёнка» в рамках национального проекта «Образование» Региональный модельный центр Свердловской области является неотъемлемым элементом образовательной экосистемы дополнительного образования Дворца молодёжи, региона, России.
Его главная задача – вывести качество дополнительного образования на новый уровень за счёт обновления содержания программ, повышения уровня подготовки, квалификации и профессионализма руководящих и педагогических работников, внедрения новых управленческих и финансовых механизмов, интеграции урочной и внеурочной деятельности, общего, профессионального и дополнительного образования.
– Мы создаём новое образовательное пространство вместе с командой единомышленников и партнёров. В нашем сообществе – Детские технопарки «Кванториум» и Центры цифрового образования «IT-куб», Фонд поддержки талантливых детей и молодёжи «Золотое сечение», все РМЦ России, Фонд развития новых форм развития. В тесном сотрудничестве мы обучаем педагогов, и те, кто прошел обучение, сами становятся преподавателями и наставниками. Формируем родительское сообщество как равноправных партнеров образовательного процесса и становимся своеобразным центром управления полётами, – рассказала Екатерина Корьякина, директор Регионального модельного центра Дворца молодёжи.
В 2020 году РМЦ начал обучение по программам повышения квалификации, профессиональной переподготовки и профессионального обучения. «Все программы направлены на подготовку педагога нового формата – двигателя процесса изменений. Наши программы уникальны – все они носят практикоориентированный характер и завершаются защитой индивидуальных или групповых проектов», – подчеркнула Екатерина Александровна.
За время своего существования Центр внедрил систему персонифицированного финансирования дополнительного образования (ПФДО) в регионе – сертифицировал более 1000 программ дополнительного образования, в том числе для одарённых детей и детей с ОВЗ. Благодаря работе РМЦ в Свердловской области было выдано более 230 тысяч сертификатов ПФДО.
Для решения задач дополнительного образования по выявлению, развитию и поддержке одарённых и талантливых детей РМЦ сотрудничает с Фондом «Золотое сечение». Так, в этом году было проведено обучение вожатых на сменах Фонда в загородном образовательном центре «Таватуй», а в ноябре там пройдут курсы повышения квалификации для педагогов по работе с талантливыми детьми.
По словам Екатерины Корьякиной, ответом на вызовы системе образования в регионе станут сценарии и модели развития дополнительного образования, учитывающие специфику каждого муниципалитета. А для того, чтобы они были эффективными, РМЦ привлекает к работе специалистов Муниципальных образовательных центров, Управлений образования, использует опыт других регионов и специалистов разных сфер деятельности.
Компания «Вектор», Воронеж: адрес, телефон, отзывы — Информационно
Магазин — салон «Вектор-Керамика» — движение в нужном направлении. Обратившись в салон-магазин Вектор-Керамика, Вы сделали правильный выбор! Мы поможем Вам создать неповторимый интерьер, объединив вместе стиль, комфорт, красоту, долговечность и при этом не переплачивать лишних денег.
Не секрет, что выбор керамической плитки, панно, декоров и мозаики может стать утомительным. А между тем выбор плитки — один из самых ответственных этапов в ремонте, ведь она выбирается, как правило, на многие годы. Поэтому должна сочетать в себе красоту, прочность, долговечность и лёгкость в уходе. Чтобы сделать Ваш выбор как можно более лёгким и приятным, мы постарались представить в нашем магазине как можно больше коллекций самых различных стилей и направлений.
В магазине «Вектор-Керамика» Вы найдёте лучшие коллекции керамической плитки, керамогранита самых известных фабрик Европы, России, Украины, Белоруссии и Китая. Мы работаем только с производителями, хорошо зарекомендовавшими себя и подтверждающими свою репутацию в течение многих лет. В магазине «Вектор-Керамика» Вы найдёте огромное количество коллекций керамической плитки и керамогранита, разнообразных по стилю, назначению и цене. К тому же, широчайший ассортимент постоянно обновляется и дополняется интересными новинками. Специализированные магазины — салоны керамической плитки не оставляют наших покупателей равнодушными. Приветливые продавцы вместе с дизайнерами — помогают и в выборе плитки и грамотном оформлении интерьера.
Мы работаем только с производителями, хорошо зарекомендовавшими себя и подтверждающими свою репутацию в течение многих лет. В магазине «Вектор-Керамика» Вы найдёте огромное количество коллекций керамической плитки и керамогранита, разнообразных по стилю, назначению и цене. К тому же, широчайший ассортимент постоянно обновляется и дополняется интересными новинками. Специализированные магазины — салоны керамической плитки не оставляют наших покупателей равнодушными. Приветливые продавцы вместе с дизайнерами — помогают и в выборе плитки и грамотном оформлении интерьера.
Индивидуальные дизайны в формате 3D радуют нашего клиента, определяют его в выборе собственного уникального стиля. Огромный ассортимент ведущих производителей Италии, Испании, России, Украины, Белоруссии, Польши и др. Компания «Вектор-Керамика» уже более 15 лет на Российском рынке. Мы продаем керамическую плитку оптом и в розницу.
С 01.112017 г. Компания «Вектор-Керамика» объединилась с компанией «Золотое сечение».
ОСНОВЫ КОМПОЗИЦИИ: Золотое сечение — PhotoDzen.com
15 Апреля 2015
Композиция — это распределение предметов и фигур в пространстве, установление соотношения объёмов, света и тени, цвета и т. п. Существуют разные способы и правила для создания гармоничной композиции. Когда мы смотрим вокруг невооруженным глазом, наш мозг быстро выхватывает интересные сюжеты, объекты. Камера же фиксирует все подряд. Поэтому это ваша задача выбрать основной объект, сделать центром внимания в кадре, а окружающие его другие объекты превратить в фон или сделать частью истории, которую Вы хотите «рассказать» своим снимком.
Фотографии с правильно выбранной композицией заставляют задержать на них взгляд и рассмотреть детали. Они рассказывают историю, создают настроение и заставляют задуматься.
Золотое сечение в фотографии — основной и мощный инструмент для получения динамичных, интересных снимков. Правило золотого сечения встречается и в природе, причем повсеместно. О нем знали еще в древнем Египте. Пропорции пирамиды Хеопса, храмов, барельефов, предметов быта и украшений из гробницы Тутанхамона свидетельствуют, что египетские мастера пользовались соотношениями золотого сечения при их создании. В фасаде древнегреческого храма Парфенона также присутствуют золотые пропорции. Этот феномен изучил и стал применять на практике известный ученый, художник и скульптор Леонардо Да Винчи.
Для желающих узнать больше — видео:
Мы затронем только практическую часть применения золотого сечения в фотографии. Кадр условно делиться на три части по горизонтали и вертикали:
При пересечении горизонтальной и вертикальной линии образуется особая точка – «точка силы» или «узел внимания».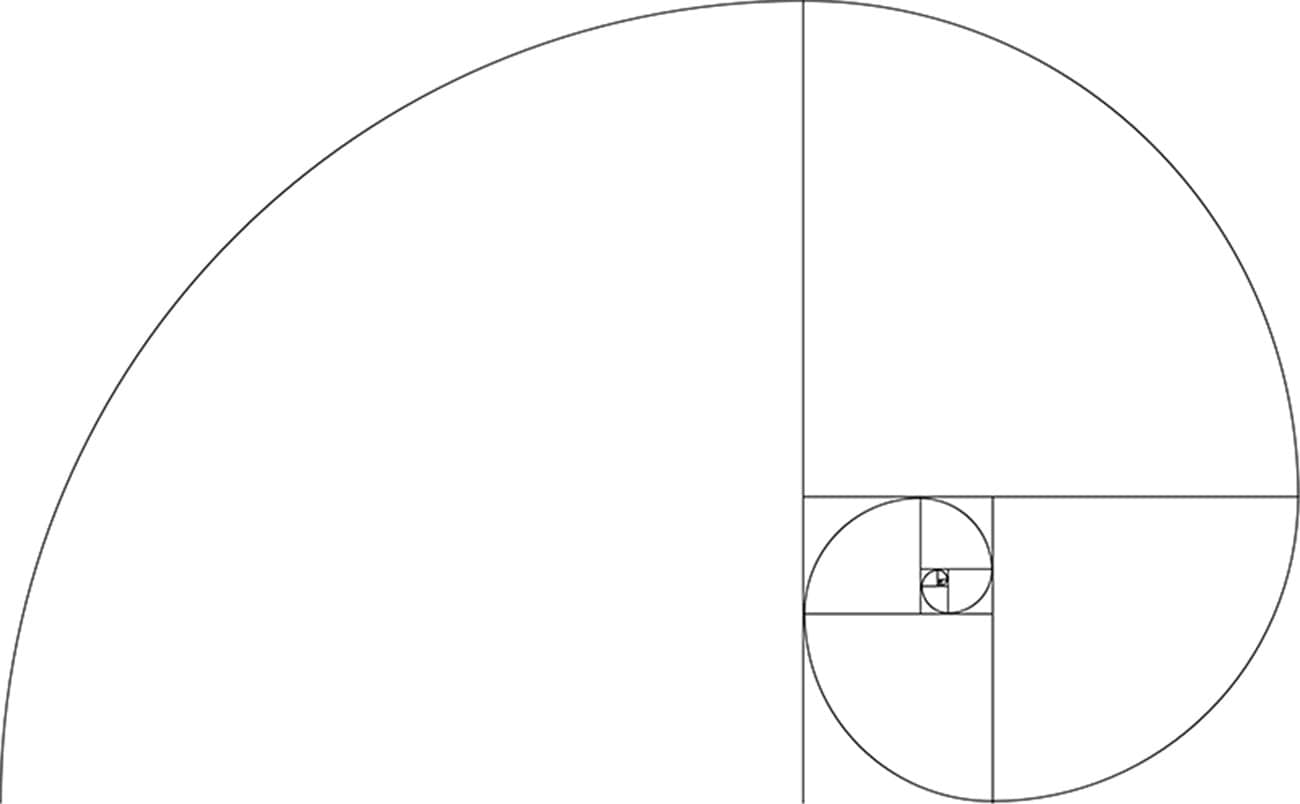 Их четыре – именно в этих точках лучше располагать главные объекты кадра, именно на них останавливается взгляд в независимости от формата кадра или картины.
Их четыре – именно в этих точках лучше располагать главные объекты кадра, именно на них останавливается взгляд в независимости от формата кадра или картины.
Практические советы:
- Если расположитьлинию горизонта по одной из горизонтальных линий, то кадр будет смотреться гармоничней. Но на какой из линий, на верхней или нижней?
- Если вы хотите сконцентрировать внимание зрителя на земле или воде – то лучше на верхней.
- Если же вы делаете акцент на интересном, выразительном небе – то на нижней.
- Если вы снимаете портрет, то лучше размещать глаза на верхней горизонтальной линии.
- Если вы снимаете человека в полный рост, то лучше размещать его на правой или левой вертикальной линии.
- Очень важно следить за тем, в какую сторону направляется человек, или куда направлен его взгляд.
 Например, если человек смотрит влево, то его соответственно необходимо разместить на правой горизонтальной линии, что бы перед ним оставалось пространство.
Например, если человек смотрит влево, то его соответственно необходимо разместить на правой горизонтальной линии, что бы перед ним оставалось пространство.
В современных фотоаппаратах уже есть функция-подсказка, которая отображает линии правила третей на мониторе или в видоискателе.
Размещайте важные части композиции вдоль линий, а самое главное — на их пересечении.
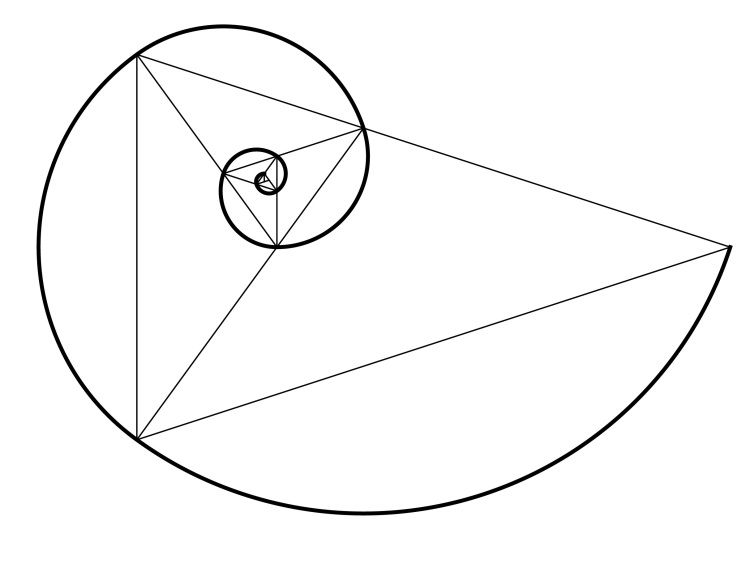
Золотое сечение может прослеживаться не только в прямоугольной сетке, но и в диагоналях или спирали. Принцип расположения предметов – тот же, по основным линиям и в точках их пересечений.
Фото: Steve McCurry
Фото: Joe McNally
Фото: Richard Barnabe
ДИАГОНАЛЬНОЕ ЗОЛОТОЕ СЕЧЕНИЕ
Применяя правило золотого сечения проводим диагонали и получаем прямоугольник состоящий из трех секторов. Этот прямоугольник можно поворачивать как угодно Если скомпонуете свой кадр так, чтобы три разных объекта примерно располагались в этих секторах, а главные объекты в более крупных секциях — то композиция будет выглядеть очень гармонично.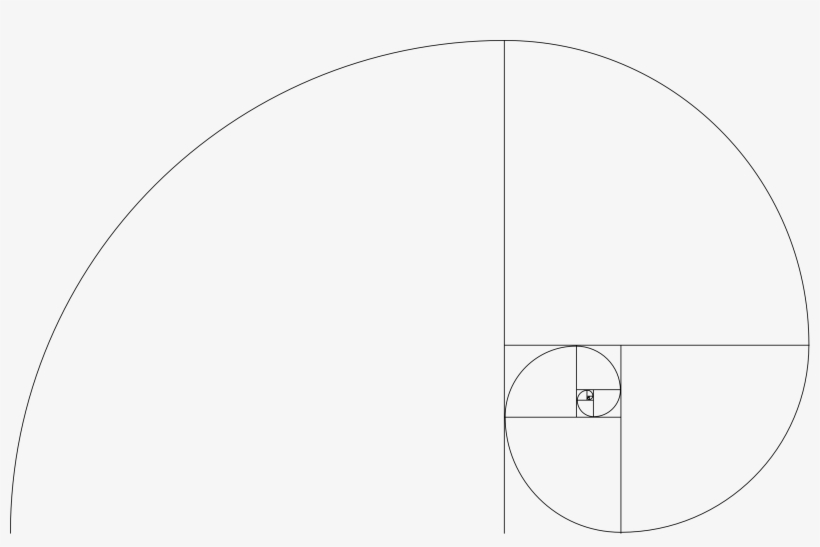
Это правило используется, если у вас в кадре есть несколько областей, различающихся по смыслу.
СПИРАЛЬНОЕ ЗОЛОТОЕ СЕЧЕНИЕ
Спирали очень распространены в природе. Форма спирально завитой раковины привлекла внимание Архимеда. Он изучал ее и вывел уравнение спирали. Спираль, вычерченная по этому уравнению, называется его именем. Увеличение ее шага всегда равномерно. В настоящее время спираль Архимеда широко применяется в технике. Гёте называл спираль — «кривой жизни».
Используя эту спираль при построении композиции в кадре (ее можно перевернуть вверх ногами или в другую сторону), мы получим кадр с четко выраженным предметом в центре спирали.
Фото: Джона Лемьё (John Lemieux)
Фото: Анри Картье-Брессон
Больше фотографируйте и экспериментируйте. Удачи!
Другие статьи о композиции в фотографии:
Золотое сечение в фотографии
Сила линий в фотографии
Формат кадра в фотографии
Контраст в фотографии (Часть 1)
Контраст в фотографии (Часть 2)
Точка съемки и выбор плана
Ракурс
Геометрия в фотографии
Равновесие
Файл:Спираль Фибоначчи 34.
 svg — Wikimedia Commons
svg — Wikimedia CommonsРезюме[править]
| ОписаниеСпираль Фибоначчи 34.svg | Английский: Спираль Фибоначчи с размером квадрата до 34 |
| Дата | 17 марта 2008 г. |
| Источник | Собственная работа с использованием: Inkscape |
| Автор | Пользователь: Диклион |
Это векторное изображение было создано с помощью Inkscape.
Лицензирование[править]
| Общественное достояниеОбщественное достояниеfalsefalse |
| Я, владелец авторских прав на это произведение, делаю это произведение общественным достоянием . Это применимо во всем мире. В некоторых странах это может быть невозможно по закону; если да: Я предоставляю любому право использовать это произведение в любых целях без каких-либо условий, если только такие условия не требуются по закону.  |
«Умная» рекурсивная версия[править]
Эта версия зависит от SVG 1.2 и пока плохо отображается в rsvg (00:25, 23 апреля 2013 г. (UTC)).
Щелкните дату/время, чтобы просмотреть файл в том виде, в котором он был в то время.
| Дата / время | Thumbnail | Размеры | User | Комментарий | |||
|---|---|---|---|---|---|---|---|
| Ток | 22:19, 22 апреля 2013 | 22:19, 22 апреля 2013 | 9000 × 579 (1 Kb) | YSangkok ( Talk | вклад) | RSVG не поддерживает стилы, используют группы | ||
| 22:13, 22 апреля 2013 | 915 × 579 (1 кБ) | ysangkok (Talk | Place) | SCOUR | ||||
| 9000 × 579 (7 кб) | (7 кб) | Dicklyon (Talk | Placts) | {{информация | Описание = Фибоначчи Спираль с квадратными размерами до 34 |Источник=нарисовано в Inkscape |Дата=17 марта 2008 |Автор= |Разрешение= |other_versions= }} |
Этот файл нельзя перезаписать.
Следующие 2 страницы используют этот файл:
- Спираль
- Файл:BiomineralizationCyclicRhythms Ingrid.svg
Этот файл содержит дополнительную информацию, такую как метаданные Exif, которые могли быть добавлены цифровой камерой, сканером или программой, используемой для его создания или оцифровки. Если файл был изменен по сравнению с исходным состоянием, некоторые детали, такие как метка времени, могут не полностью отражать детали исходного файла. Временная метка настолько точна, насколько точны часы в камере, и она может быть совершенно неправильной.
Пи и золотое сечение
Два моих любимых числа — число пи:
.и золотое сечение:
Они родственники:
Грег Иган и я придумали эту формулу на прошлых выходных. Вероятно, это не ново и, конечно, не удивит экспертов, но все равно интересно придумать такую формулу. Позвольте мне объяснить, как мы это сделали.
История имеет фрактальную текстуру. Это не совсем самоподобный , но чем ближе вы смотрите на любой инцидент, тем больше мелких деталей вы видите.Упрощенные рассказы об истории математики и физики, которые мы изучаем в школе, подобны расплывчатым изображениям множества Мандельброта. Вы можете видеть общую форму, но действительно интересные вещи скрыты.
Это не совсем самоподобный , но чем ближе вы смотрите на любой инцидент, тем больше мелких деталей вы видите.Упрощенные рассказы об истории математики и физики, которые мы изучаем в школе, подобны расплывчатым изображениям множества Мандельброта. Вы можете видеть общую форму, но действительно интересные вещи скрыты.
Франсуа Виет — французский математик, которого нет в этих упрощенных историях. Он изучал право в Пуатье, который окончил в 1559 году. Он начал свою карьеру в качестве поверенного на довольно высоком уровне, занимаясь делами вдовы короля Франции Франциска I, а также Марии, королевы Шотландии.Но его истинным интересом всегда была математика. Друг сказал, что может думать об одном вопросе до трех дней, положив локоть на стол, питаясь, не меняя положения.
Тем не менее, он очень преуспел в юриспруденции. К 1590 году он работал на короля Генриха IV. Король восхищался его математическими способностями, и Виет вскоре подтвердил свою ценность, взломав испанский шифр, что позволило французам прочитать все испанские сообщения, которые они смогли получить.
В 1591 году Франсуа Виет выпустил важную книгу, в которой представил так называемую новую алгебру: символический метод работы с полиномиальными уравнениями.Это заслуживает большей известности; он был хорошо знаком Декарту и другим, и он был важным предшественником наших современных обозначений и методов. Например, он подчеркивал осторожность при использовании переменных и выступал за обозначение известных величин согласными, а неизвестных величин — гласными. (Позже люди перешли на использование букв в начале алфавита для известных величин и букв в конце, как для неизвестных.)
В 1593 году он выпустил еще одну книгу, Variorum De Rebus Mathematicis Responsorum, Liber VIII .Среди прочего, он включает в себя формулу для числа пи. В модернизированных обозначениях это выглядит так:
Это замечательно! Во-первых, это выглядит круто. Во-вторых, это самый ранний известный пример бесконечного произведения в математике. В-третьих, это самая ранняя известная формула для точного значения числа пи. На самом деле это самая ранняя формула, представляющая число как результат бесконечного процесса, а не конечного вычисления! Итак, формулу Виете называют началом анализа.В своей статье «Жизнь числа Пи» Джонатан Борвейн пошел еще дальше и назвал формулу Виета «рассветом современной математики».
На самом деле это самая ранняя формула, представляющая число как результат бесконечного процесса, а не конечного вычисления! Итак, формулу Виете называют началом анализа.В своей статье «Жизнь числа Пи» Джонатан Борвейн пошел еще дальше и назвал формулу Виета «рассветом современной математики».
Как Виете придумал свою формулу? Я не читал его книгу, но идея кажется довольно ясной. Площадь единичного круга равна пи. Итак, вы можете все лучше и лучше аппроксимировать число пи, вычисляя площадь квадрата, вписанного в этот круг, затем восьмиугольника, затем 16-угольника и так далее:
Если вы подсчитаете эти площади умным способом, вы получите следующий ряд чисел:
и так далее, где площадь правильного n -угольника, вписанного в единичную окружность.Таким образом, Виет сделал лишь небольшой шаг (но бесконечный скачок для человечества), чтобы прийти к заключению, что
или, если квадратные корни в знаменателе вас не устраивают:
Основная идея здесь не удивила бы Архимеда, который строго доказал, что
путем аппроксимации длины окружности с помощью обычного 96-угольника. Так как вы можете нарисовать правильный 96-угольник с помощью линейки и циркуля, взяв равносторонний треугольник и разделив его края пополам, чтобы получить шестиугольник, разделив его пополам, чтобы получить 12-угольник, и так далее до 96.В более современном образе мышления вы можете выяснить все, что вам нужно знать, начав с угла и используя формулы половинного угла 4 раза, чтобы вычислить синус или косинус . И действительно, до того, как появился Виет, Людольф ван Сеулен вычислил число пи до 35 цифр, используя правильный многоугольник со сторонами! Итак, смелая новая идея Виета состояла в том, чтобы дать точную формулу для числа пи, которая включала бы бесконечный процесс .
Так как вы можете нарисовать правильный 96-угольник с помощью линейки и циркуля, взяв равносторонний треугольник и разделив его края пополам, чтобы получить шестиугольник, разделив его пополам, чтобы получить 12-угольник, и так далее до 96.В более современном образе мышления вы можете выяснить все, что вам нужно знать, начав с угла и используя формулы половинного угла 4 раза, чтобы вычислить синус или косинус . И действительно, до того, как появился Виет, Людольф ван Сеулен вычислил число пи до 35 цифр, используя правильный многоугольник со сторонами! Итак, смелая новая идея Виета состояла в том, чтобы дать точную формулу для числа пи, которая включала бы бесконечный процесс .
Теперь давайте подробно рассмотрим, как работает формула Виете.Поскольку нет необходимости начинать с квадрата, мы могли бы также начать с обычного n -угольника, вписанного в круг, и многократно делить его стороны пополам, получая все более и более лучшие приближения к числу пи. Если мы начнем с пятиугольника, то получим формулу для числа пи, включающую золотое сечение!
Если мы начнем с пятиугольника, то получим формулу для числа пи, включающую золотое сечение!
У нас есть
, поэтому мы также можем вычислить число пи, начав с обычного n -угольника и многократно удваивая количество вершин:
Ключевой прием — написать как «телескопический продукт»:
Таким образом, принимая предел, мы получаем
, где мы начинаем с площади n -угольника и продолжаем «корректировать» его, чтобы получить площадь 2n -угольника, 4n -угольника, 8n -угольника и так далее.
Существует простая формула площади правильного n -угольника, вписанного в окружность. Вы можете разбить его на прямоугольные треугольники, каждый из которых имеет основание и высоту и, таким образом, площадь:
Таким образом,
Это позволяет нам понять, как изменяется площадь, когда мы удваиваем количество вершин:
Это красиво и просто, но нам действительно нужна рекурсивная формула для этой величины. Давайте определим
Давайте определим
Почему коэффициент 2? Это несколько упрощает наши расчеты.Мы можем выразить в терминах использования формулы половинного угла для косинуса:
Теперь мы готовы повеселиться! У нас есть
поэтому, используя нашу рекурсивную формулу, которая справедлива для любого , мы получаем
Я думаю, что это заслуживает того, чтобы называться обобщенной формулой Виете . И действительно, если начать с квадрата, то получится
.и
дает формулу Виета:
по желанию!
А что, если мы начнем с пятиугольника? Для этого полезно вспомнить красивый, но немного неясный триггерный факт:
.и чуть менее красивый:
Это легко доказать, и я покажу вам, как позже.Пока обратите внимание, что они подразумевают
и
Таким образом, формула
дает нам
или, немного очистив его, формула, которую мы хотим:
Вуаля!
Можно еще многое сказать, но позвольте мне просто объяснить немного неясные факты тригонометрии, которые нам нужны.![]() Чтобы вывести их, я считаю полезным вспомнить, что правильный пятиугольник и пентаграмма внутри него содержат множество подобных треугольников:
Чтобы вывести их, я считаю полезным вспомнить, что правильный пятиугольник и пентаграмма внутри него содержат множество подобных треугольников:
Используя тот факт, что все эти треугольники подобны, легко показать, что для любого из них отношение длинной стороны к короткой равно 1, так как
Другим важным фактом является то, что пентаграмма делит внутренний угол правильного пятиугольника на три части, разбивая внутренний угол на 3 угла:
Опять же, это легко и весело показать.
Комбинируя эти факты, мы можем доказать, что
и
Чтобы доказать первое уравнение, разрежьте один из этих золотых треугольников на два прямоугольных треугольника и сделайте то, чему научились в старшей школе. Чтобы доказать второе, проделайте то же самое с одним из коротких равнобедренных треугольников в приседе:
Исходя из этих уравнений и используя , мы можем показать
и, просто для полноты (здесь он нам не нужен):
Для этого требуются слегка раздражающие вычисления, где помогает использование идентификатора
.
Ладно, пока все! Но если вы хотите повеселиться, попробуйте решить пару головоломок:
.Головоломка 1. Мы получили формулы для числа пи, начиная с квадрата или правильного пятиугольника. Какую формулу вы получите, исходя из равностороннего треугольника?
Задача 2. Используя обобщенную формулу Виета, докажите формулу Эйлера
И наоборот, используйте формулу Эйлера для доказательства обобщенной формулы Виете.
Итак, можно сказать, что реальная точка формулы Виета и ее обобщенной версии — это не какое-то особое свойство числа пи, а формула Эйлера.
Эта запись была опубликована во вторник, 7 марта 2017 г., в 16:47 и находится в разделе «Математика».Вы можете следить за любыми ответами на эту запись через ленту RSS 2.0. Вы можете оставить отзыв или вернуться со своего сайта.
Почтовая навигация
» Предыдущий пост Следующее сообщение »Эффективное и безошибочное флуоресцентное мечение генов в органоидах человека без расщепления двухцепочечной ДНК
Abstract
CRISPR-ассоциированные нуклеазы являются мощными инструментами для точного редактирования генома модельных систем, включая органоиды человека. Текущие методы, описывающие мечение флуоресцентных генов в органоидах, основаны на генерации двухцепочечных разрывов ДНК (DSB) для стимуляции опосредованной гомологии репарации (HDR) или негомологичного соединения концов (NHEJ) интеграции желаемого нокаута. Основным недостатком, связанным с редактированием генома, опосредованным DSB, является необходимая клональная селекция и экспансия органоидов-кандидатов для проверки геномной целостности целевого локуса и подтверждения отсутствия нецелевых вставок. Напротив, одновременный разрыв геномного локуса и вектора-мишени, известный как in-trans-paired nicking (ITPN), стимулирует эффективное HDR-опосредованное редактирование генома для создания больших нокаутов без введения DSB.Здесь мы показываем, что ITPN позволяет быстро, высокоэффективно и без делеций флуоресцентно маркировать гены в нормальных и раковых органоидах человека. Подчеркивая простоту и эффективность ITPN, мы создаем органоиды с тройным флуоресцентным нокаутом, в которых 3 геномных локуса были одновременно модифицированы в одном раунде нацеливания.
Текущие методы, описывающие мечение флуоресцентных генов в органоидах, основаны на генерации двухцепочечных разрывов ДНК (DSB) для стимуляции опосредованной гомологии репарации (HDR) или негомологичного соединения концов (NHEJ) интеграции желаемого нокаута. Основным недостатком, связанным с редактированием генома, опосредованным DSB, является необходимая клональная селекция и экспансия органоидов-кандидатов для проверки геномной целостности целевого локуса и подтверждения отсутствия нецелевых вставок. Напротив, одновременный разрыв геномного локуса и вектора-мишени, известный как in-trans-paired nicking (ITPN), стимулирует эффективное HDR-опосредованное редактирование генома для создания больших нокаутов без введения DSB.Здесь мы показываем, что ITPN позволяет быстро, высокоэффективно и без делеций флуоресцентно маркировать гены в нормальных и раковых органоидах человека. Подчеркивая простоту и эффективность ITPN, мы создаем органоиды с тройным флуоресцентным нокаутом, в которых 3 геномных локуса были одновременно модифицированы в одном раунде нацеливания. Кроме того, мы создали модельные системы с аллель-специфическими показаниями путем дифференциальной модификации материнских и отцовских аллелей за один шаг. ITPN с использованием нашей палитры целевых векторов, общедоступной от Addgene, идеально подходит для создания безошибочных гетерозиготных нокаутов в человеческих органоидах.
Кроме того, мы создали модельные системы с аллель-специфическими показаниями путем дифференциальной модификации материнских и отцовских аллелей за один шаг. ITPN с использованием нашей палитры целевых векторов, общедоступной от Addgene, идеально подходит для создания безошибочных гетерозиготных нокаутов в человеческих органоидах.
Образец цитирования: Bollen Y, Hageman JH, van Leenen P, Derks LLM, Ponsioen B, Buissant des Amorie JR, et al. (2022)Эффективное и безошибочное мечение флуоресцентных генов в органоидах человека без расщепления двухцепочечной ДНК. PLoS Биол 20(1): е3001527. https://doi.org/10.1371/journal.pbio.3001527
Академический редактор: Мэдлин Ланкастер, СОЕДИНЕННОЕ КОРОЛЕВСТВО
Получено: 10 мая 2021 г.; Принято: 5 января 2022 г .; Опубликовано: 28 января 2022 г.
Авторское право: © 2022 Bollen et al. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Авторы подтверждают, что все данные, лежащие в основе выводов, полностью доступны без ограничений. Все соответствующие данные содержатся в документе и файлах со вспомогательной информацией. Все алгоритмы, использованные для отображения [gatk.wideinstitute.org], вызов мутаций [https://github.com/ToolsVanBox/NF-IAP] и фильтрация мутаций [https://github.com/ToolsVanBox/SMuRF, https://github.com/hartwigmedical/ gridss-purple-linx] общедоступны. Необработанные файлы FCS доступны в базе данных FlowRepository (flowrepository.org) и доступны с использованием идентификатора репозитория FR-FCM-Z4PJ.
Финансирование: Эта работа является частью Института Онкод, который частично финансируется Голландским онкологическим обществом.![]() HGJS получил стартовый грант Европейского исследовательского совета (ERC) (IntratumoralNiche), номер проекта 803608 (https://erc.europa.eu/funding/starting-grants) и NWO TOP. YB был поддержан стратегическим альянсом между Университетом Твенте и UMC Utrecht в области передового биопроизводства (до LWMMT и HJGS). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
HGJS получил стартовый грант Европейского исследовательского совета (ERC) (IntratumoralNiche), номер проекта 803608 (https://erc.europa.eu/funding/starting-grants) и NWO TOP. YB был поддержан стратегическим альянсом между Университетом Твенте и UMC Utrecht в области передового биопроизводства (до LWMMT и HJGS). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Сокращения: ДСБ, двухцепочечный разрыв; HDR, репарация, направленная по гомологии; ИТПК, ин-транс парное расщепление; ИТПН, внутритранс-парные ники; ММЕЖ, соединение концов, опосредованное микрогомологией; NHEJ, негомологичное соединение концов; ВГС, полногеномное секвенирование
Введение
С момента разработки эффективной технологии редактирования генома молекулярные и клеточные биологические исследования все больше полагаются на генетически модифицированные модельные системы in vitro. В частности, визуализация эндогенных белков с помощью флуоресцентных нокаут-репортеров позволяет точно оценить их внутриклеточную локализацию и динамику при клеточном гомеостазе и заболевании [1].
В частности, визуализация эндогенных белков с помощью флуоресцентных нокаут-репортеров позволяет точно оценить их внутриклеточную локализацию и динамику при клеточном гомеостазе и заболевании [1].
Органоиды, в частности человеческого происхождения, представляют собой модельные системы следующего поколения, которые воспроизводят архитектуру и функциональность тканей in vivo более точно, чем двумерные клеточные линии [2]. Тем не менее, точная разработка больших репортеров с нокаутом в органоидах может быть трудоемкой при использовании обычных стратегий, опосредованных CRISPR, для стимуляции редактирования, основанного на гомологичном восстановлении (HDR) [3–5] или негомологичном соединении концов (NHEJ) [6].Хотя в целом эти стратегии эффективны, они основаны на генерации геномных двухцепочечных разрывов (DSB) с помощью CRISPR-ассоциированных нуклеаз, что часто приводит к индел-мутациям как на, так и вне мишени в результате склонной к ошибкам репарации с помощью повторяющихся циклов репарации. NHEJ. Целевые вставки часто генерируются в «нецелевом аллеле», который не несет нок-ин, и могут приводить к миссенс-мутациям или нонсенс-мутациям. Кроме того, в то время как HDR обычно приводит к безошибочной репарации, создание нокаунов посредством лигирования линеаризованного фрагмента ДНК на основе NHEJ часто приводит к вставкам в восходящих и нисходящих соединениях нокаутируемого аллеля [6,7].Следовательно, существующие протоколы нокаута по своей сути требуют проверки последовательности индивидуально отобранных клонов органоидов, что является трудоемким, трудоемким и устраняет генетическую гетерогенность в моделях органоидов, полученных из опухоли.
NHEJ. Целевые вставки часто генерируются в «нецелевом аллеле», который не несет нок-ин, и могут приводить к миссенс-мутациям или нонсенс-мутациям. Кроме того, в то время как HDR обычно приводит к безошибочной репарации, создание нокаунов посредством лигирования линеаризованного фрагмента ДНК на основе NHEJ часто приводит к вставкам в восходящих и нисходящих соединениях нокаутируемого аллеля [6,7].Следовательно, существующие протоколы нокаута по своей сути требуют проверки последовательности индивидуально отобранных клонов органоидов, что является трудоемким, трудоемким и устраняет генетическую гетерогенность в моделях органоидов, полученных из опухоли.
В клеточных линиях были получены крупные нокины без введения DSB с использованием частично инактивированного варианта никазы Cas9 D10A [8–10], который генерирует одноцепочечные разрывы ДНК (никеры) в геномной цепи, которая гибридизуется с направляющей РНК. [11].Путем одновременного разрыва геномного локуса-мишени и концов обоих плеч гомологии внутри нацеливающего вектора, стратегии, известной как in-trans-paired nicking (ITPN) [8], можно генерировать эффективные нокаут-аллели без двухцепочечного расщепления ДНК. В отличие от обычного редактирования генома, опосредованного CRISPR/Cas9, ITPN модифицирует целевые локусы с высокой точностью, поскольку одиночные геномные разрывы редко бывают мутагенными [8,12]. Избегая расщепления двухцепочечной ДНК, ITPN позволяет вставлять гетерозиготные репортеры или патогенные (зародышевые) мутации с интактными «нецелевыми» вторичными аллелями и с минимальным риском нецелевых инделей.Следовательно, нокаутированные клетки могут быть объединены, чтобы ускорить распространение и, таким образом, «время генерации» нокаутированной линии (2 недели). Объединение успешно нацеленных органоидов особенно полезно для моделей органоидов, где клональный отбор трудоемок. Более того, избегая клональной селекции, существовавшее ранее генетическое разнообразие в линиях органоидов, происходящих из опухоли, в значительной степени сохраняется. Здесь мы исследуем эффективность и точность мечения флуоресцентных генов с помощью ITPN в органоидах человека. Кроме того, мы представляем палитру простых в использовании векторных основ и протоколов для маркировки N- или C-концевых флуоресцентных генов с использованием ITPN.
В отличие от обычного редактирования генома, опосредованного CRISPR/Cas9, ITPN модифицирует целевые локусы с высокой точностью, поскольку одиночные геномные разрывы редко бывают мутагенными [8,12]. Избегая расщепления двухцепочечной ДНК, ITPN позволяет вставлять гетерозиготные репортеры или патогенные (зародышевые) мутации с интактными «нецелевыми» вторичными аллелями и с минимальным риском нецелевых инделей.Следовательно, нокаутированные клетки могут быть объединены, чтобы ускорить распространение и, таким образом, «время генерации» нокаутированной линии (2 недели). Объединение успешно нацеленных органоидов особенно полезно для моделей органоидов, где клональный отбор трудоемок. Более того, избегая клональной селекции, существовавшее ранее генетическое разнообразие в линиях органоидов, происходящих из опухоли, в значительной степени сохраняется. Здесь мы исследуем эффективность и точность мечения флуоресцентных генов с помощью ITPN в органоидах человека. Кроме того, мы представляем палитру простых в использовании векторных основ и протоколов для маркировки N- или C-концевых флуоресцентных генов с использованием ITPN.![]()
Результаты
Чтобы исследовать эффективность флуоресцентных нокаунов в органоидах человека (рис. 1А), мы разработали N-концевой нокаут mScarlet в локусе человека SEC61B . Мы построили различные векторы нацеливания, чтобы сравнить эффективность редактирования различных стратегий нокаута (рис. 1B). Чтобы стимулировать редактирование посредством NHEJ-опосредованного лигирования линеаризованного фрагмента, кодирующего mScarlet, в геномный DSB, сгенерированный Cas9 [6,7], мы сконструировали вектор, несущий последовательность, кодирующую mScarlet, с флангами копий геномного сайта-мишени Cas9.В качестве альтернативы мы включили микрогомологию 20 п.н., чтобы стимулировать геномную интеграцию через путь опосредованного микрогомологией соединения концов (MMEJ) [13]. Кроме того, мы создали векторы с гомологическими плечами размером 1 т.п.н. в соответствии с традиционной конструкцией нацеливающего вектора, которая не имеет фланкирующих сайтов-мишеней Cas9 или с фланкирующими сайтами-мишенями Cas9 для поддержки геномной интеграции через ITPN или внутритранс-парное расщепление (ITPC) [14,15]. ].
].
Рис. 1. Флуоресцентное мечение генов в органоидах человека без расщепления двухцепочечной ДНК.
(A) Схематическое представление рабочего процесса, используемого для определения эффективности флуоресцентного нокаута в органоидах человека. Для обеспечения оптимального роста после электропорации органоиды трипсинизируют до клеточной суспензии, состоящей примерно из 5 клеточных скоплений. После электропорации клеткам дают разрастись в течение 10 дней без отбора клеток, получивших нокаутные конструкции. До анализа потока органоиды могут состоять из частичных нокаутированных популяций. Чтобы зафиксировать общую эффективность нокаута, органоидные культуры трипсинизируют до одноклеточной суспензии и анализируют поток.(B) Схематическое изображение стратегии нацеливания SEC61B . mScarlet был фланкирован гомологическими плечами, соответствующими восходящим и нисходящим последовательностям N-конца локуса SEC61B человека, кодирующего бета-субъединицу транспортного белка Sec61. Cas9 был нацелен близко к началу кодирующей области с использованием гРНК, как указано (зеленая стрелка). Указаны сайты расщепления Cas9 (треугольники) и мотивы, прилегающие к протоспейсеру (черная полоса). Гомология вверх и вниз по течению представлена синим и красным цветом соответственно.Указаны композиции нацеливающих векторов, поддерживающих различные стратегии нокаута. (C) Эффективность нок-ина mScarlet в локусе SEC61B человека в модели органоида опухоли, полученной от пациента, с использованием различных стратегий нок-ина. Никазу WT или D10A (Nick) SpCas9 совместно доставляли с нацеливающими векторами, указанными в (B). Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Эффективность редактирования (% mScarlet + клеток) определяли анализом потока отдельных клеток через 10 дней после электропорации ( n = 3 независимых эксперимента).* p < 0,05 в парном тесте Ratio t .
Cas9 был нацелен близко к началу кодирующей области с использованием гРНК, как указано (зеленая стрелка). Указаны сайты расщепления Cas9 (треугольники) и мотивы, прилегающие к протоспейсеру (черная полоса). Гомология вверх и вниз по течению представлена синим и красным цветом соответственно.Указаны композиции нацеливающих векторов, поддерживающих различные стратегии нокаута. (C) Эффективность нок-ина mScarlet в локусе SEC61B человека в модели органоида опухоли, полученной от пациента, с использованием различных стратегий нок-ина. Никазу WT или D10A (Nick) SpCas9 совместно доставляли с нацеливающими векторами, указанными в (B). Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Эффективность редактирования (% mScarlet + клеток) определяли анализом потока отдельных клеток через 10 дней после электропорации ( n = 3 независимых эксперимента).* p < 0,05 в парном тесте Ratio t . Столбики погрешностей указывают SEM. На вставке показаны репрезентативные кадры локализации mScarlet- SEC61B в опухолевых органоидах пациента (масштабная линейка = 10 мкм). Базовые данные представлены в S1 Data. (D) Эффективность нок-ина mScarlet в локусе SEC61B человека в опухолевых органоидах с использованием нацеливающих векторов с разной длиной плеча гомологии, фланкированных сайтами-мишенями Cas9 и совместно доставляемых с никазой Cas9 D10A для поддержки ITPN ( n = 3 независимых эксперимента) .Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. (E) Как и в (B), схема, показывающая стратегию нацеливания для ITPN-опосредованной интеграции mScarlet (0,7 kb) или mScarlet-P2A-Puromycin R (1,4 kb) на С-конце человеческого локуса HIST1h3BC , кодирующий гистон h3B тип-1C. (F) Нокаутирующая эффективность mScarlet (mSC; 0,7 kb) или mScarlet-P2A-Puromycin R (mSC-P R ; 1.
Столбики погрешностей указывают SEM. На вставке показаны репрезентативные кадры локализации mScarlet- SEC61B в опухолевых органоидах пациента (масштабная линейка = 10 мкм). Базовые данные представлены в S1 Data. (D) Эффективность нок-ина mScarlet в локусе SEC61B человека в опухолевых органоидах с использованием нацеливающих векторов с разной длиной плеча гомологии, фланкированных сайтами-мишенями Cas9 и совместно доставляемых с никазой Cas9 D10A для поддержки ITPN ( n = 3 независимых эксперимента) .Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. (E) Как и в (B), схема, показывающая стратегию нацеливания для ITPN-опосредованной интеграции mScarlet (0,7 kb) или mScarlet-P2A-Puromycin R (1,4 kb) на С-конце человеческого локуса HIST1h3BC , кодирующий гистон h3B тип-1C. (F) Нокаутирующая эффективность mScarlet (mSC; 0,7 kb) или mScarlet-P2A-Puromycin R (mSC-P R ; 1. 4 kb) в опухолевых органоидах на С-конце локуса HIST1h3BC человека ( n = 3 независимых эксперимента). Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Разница между mSC и mSC-P R была незначительной в тесте Ratio парных t . Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. (G) Эффективность нокаута mScarlet по локусу SEC61B в нормальных и опухолевых органоидах толстой кишки человека через ITPN с использованием 1 т.п.о. опухолевые органоиды соответственно).Во всех контрольных условиях нацеливающий вектор котрансфицировали направляющей, нацеленной на другой ген. Разница между нормальными и опухолевыми органоидами KI была незначительной в двустороннем непарном тесте t . Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ). ITPC, внутритранс-парное расщепление; ITPN, парный никинг в трансе; MMEJ, соединение концов, опосредованное микрогомологией; NHEJ, негомологичное соединение концов; WT, дикий тип.
4 kb) в опухолевых органоидах на С-конце локуса HIST1h3BC человека ( n = 3 независимых эксперимента). Векторы-мишени подвергали электропорации при эквимолярных соотношениях между условиями, чтобы скорректировать различия в размере векторов. Разница между mSC и mSC-P R была незначительной в тесте Ratio парных t . Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. (G) Эффективность нокаута mScarlet по локусу SEC61B в нормальных и опухолевых органоидах толстой кишки человека через ITPN с использованием 1 т.п.о. опухолевые органоиды соответственно).Во всех контрольных условиях нацеливающий вектор котрансфицировали направляющей, нацеленной на другой ген. Разница между нормальными и опухолевыми органоидами KI была незначительной в двустороннем непарном тесте t . Столбики погрешностей указывают SEM. Базовые данные представлены в S1 Data. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ). ITPC, внутритранс-парное расщепление; ITPN, парный никинг в трансе; MMEJ, соединение концов, опосредованное микрогомологией; NHEJ, негомологичное соединение концов; WT, дикий тип.
https://doi.org/10.1371/journal.pbio.3001527.g001
Векторы-мишени подвергали коэлектропорации с экспрессионными конструкциями SpCas9 никазы дикого типа или D10A в модели органоида опухоли, полученной от пациента, полученной из биобанка колоректального рака [16] . Мы визуально подтвердили ожидаемую локализацию mScarlet в нокаут-ин органоидах для каждого условия перед проточным анализом клеток mScarlet + через 10 дней после электропорации (рис. 1C). Фланкирование гомологических плеч с сайтами-мишенями Cas9 для стимуляции ITPN или ITPC привело к значительно более высокой эффективности редактирования по сравнению с традиционной конструкцией нацеливающего вектора с той же длиной гомологического плеча (рис. 1C и S1A).Кроме того, условия NHEJ и MMEJ были менее эффективными по сравнению с нацеливающими векторами с длинными гомологическими плечами, в частности, в сочетании с никазой Cas9. Примечательно, что ITPN привела к аналогичной доле нокаутированных клеток по сравнению с традиционной стратегией нокаута, в которой используется Cas9 дикого типа и нацеливающие векторы без фланкирования целевых сайтов Cas9.
Чтобы исследовать достоверность флуоресцентных нокаутов, опосредованных ITPN, мы провели анализ последовательности на поликлональных линиях нокаута, которые были получены в соответствии с описанными выше условиями.Чтобы определить риск нецелевых вставок, мы проанализировали точность вторичного аллеля, который не несет в себе нокаут, в качестве прокси для наиболее вероятных кандидатов на нецелевые модификации. Используя анализ TIDE [17], мы показываем, что условия Cas9 дикого типа приводят к высокой частоте вставок во вторичном аллеле, тогда как нокаутированные органоиды, полученные с помощью ITPN, демонстрируют> 99% целостность последовательности их вторичного аллеля (S2 Fig). Затем, чтобы исследовать достоверность нокаунов, опосредованных ITPN, мы создали 11 клональных линий нокаута из условия ITPN и исследовали аллели нокаута с помощью секвенирования по Сэнгеру.Все нокины содержали интактные 5′- и 3′-соединения, и не было обнаружено никаких доказательств тандемной интеграции (рис. S3A). Более того, в соответствии с предыдущим анализом TIDE на поликлональных культурах мы подтвердили отсутствие вставок в нецелевом аллеле гетерозиготных клонов (рис. S3B). Наконец, чтобы исключить наличие нецелевого редактирования, мы выполнили секвенирование всего генома (WGS) на 3 из 11 клональных линий нокаута, опосредованных ITPN. Мы исследовали количество соматических мутаций этих клонов в 166 областях, которые, как было предсказано in silico, вероятно, являются нецелевыми локусами протоспейсеров.Никаких геномных аберраций не было выявлено в немодифицированном аллеле, предсказанных нецелевых областях протоспейсера или 200 основаниях, окружающих предсказанные сайты (таблица S1). Отсутствие всех вариантов, начиная от замен одиночных оснований и заканчивая точками разрыва структурных вариаций, подтверждает отсутствие мутаций из-за неправильно репарированной нецелевой нуклеазной активности, а также нецелевых интеграций нокаутной кассеты.
S3A). Более того, в соответствии с предыдущим анализом TIDE на поликлональных культурах мы подтвердили отсутствие вставок в нецелевом аллеле гетерозиготных клонов (рис. S3B). Наконец, чтобы исключить наличие нецелевого редактирования, мы выполнили секвенирование всего генома (WGS) на 3 из 11 клональных линий нокаута, опосредованных ITPN. Мы исследовали количество соматических мутаций этих клонов в 166 областях, которые, как было предсказано in silico, вероятно, являются нецелевыми локусами протоспейсеров.Никаких геномных аберраций не было выявлено в немодифицированном аллеле, предсказанных нецелевых областях протоспейсера или 200 основаниях, окружающих предсказанные сайты (таблица S1). Отсутствие всех вариантов, начиная от замен одиночных оснований и заканчивая точками разрыва структурных вариаций, подтверждает отсутствие мутаций из-за неправильно репарированной нецелевой нуклеазной активности, а также нецелевых интеграций нокаутной кассеты.
В совокупности эти данные показывают, что ITPN обеспечивает высокоэффективную флуоресцентную маркировку генов без инделей в органоидах человека и делает ненужным подтверждение последовательности клональных линий. Следовательно, все нокаут-органоиды могут быть немедленно объединены, чтобы ускорить расширение отредактированной линии органоидов и сохранить генетическое разнообразие моделей опухолевых органоидов, полученных от пациентов.
Следовательно, все нокаут-органоиды могут быть немедленно объединены, чтобы ускорить расширение отредактированной линии органоидов и сохранить генетическое разнообразие моделей опухолевых органоидов, полученных от пациентов.
Традиционный дизайн векторов-мишеней требует длинных ветвей гомологии, чтобы максимизировать вероятность гомологичной рекомбинации между геномным локусом и вектором-мишенью. Однако векторы с длинными гомологическими плечами сложны для сборки и неудобны для локус-специфического генотипирования с помощью ПЦР.Чтобы исследовать, теряется ли эффективность мечения флуоресцентных генов, когда ITPN опосредуется более короткими плечами гомологии, мы создали серию векторов-мишеней с уменьшающейся гомологией. В локусе SEC61B требование гомологии для ITPN-опосредованной интеграции mScarlet достигло пика при 800 п.н. (рис. 1D и S1B). В то время как векторы с более короткими гомологическими плечами сопровождались более низкой эффективностью редактирования, их было достаточно для создания линий нокаута, и они могут быть предпочтительными в ситуациях сложной сборки векторов и/или генотипирования.
Затем, чтобы выяснить, влияет ли размер нок-ина на эффективность редактирования, мы разработали нок-ин С-конца в локусе HIST1h3BC и сконструировали векторы-мишени с гомологией 500 п.н. для интеграции либо mScarlet (0,7 т.п.н.), либо mScarlet-P2A- Puro (1,4 т.п.н.) (рис. 1E). Удивительно, но мы не обнаружили существенной разницы в эффективности нокаута между двумя вариантами нацеливающего вектора, что позволяет предположить, что размер нокаута в диапазоне <1,4 т.п.о. не оказывает заметного влияния на эффективность редактирования с помощью ITPN (рис. 1F и S1C).
Поскольку SEC61B и HIST1h3BC являются повсеместно экспрессируемыми генами, мы решили исследовать, можем ли мы ввести mScarlet-P2A-Blast в нормальные органоиды толстой кишки человека на С-конце KRT20 , который экспрессируется исключительно в дифференцированных клетки. После короткого импульса селекции бластицидином мы наблюдали клональные органоиды с субпопуляцией клеток, демонстрирующих ожидаемую цитоплазматическую красную флуоресценцию (S4 Fig). Поскольку дифференцированные клетки не образуют органоиды так же эффективно, как стволовые клетки, линии, которые включают флуоресцентные нокауты в генах дифференцировки, таких как KRT20 , лучше всего генерируются либо с использованием короткого импульса селекции, либо путем ручного отбора и объединения клональных органоидов, которые содержат (некоторые ) флуоресцентные клетки.
Поскольку дифференцированные клетки не образуют органоиды так же эффективно, как стволовые клетки, линии, которые включают флуоресцентные нокауты в генах дифференцировки, таких как KRT20 , лучше всего генерируются либо с использованием короткого импульса селекции, либо путем ручного отбора и объединения клональных органоидов, которые содержат (некоторые ) флуоресцентные клетки.
Наконец, мы сравнили эффективность N-концевого нокаута mScarlet в локусе SEC61B между опухолевыми и нормальными органоидами толстой кишки (рис. 1G и S1D). Эффективность нокаута в опухолевых органоидах была выше (хотя и незначительной), что может быть связано с разницей в условиях культивирования и эффективности электропорации.
Основным недостатком создания векторов-мишеней с гомологическими плечами, фланкированными сайтами-мишенями Cas9 на их концах, является трудоемкое молекулярное клонирование.Чтобы ускорить клонирование векторов-мишеней для мечения флуоресцентных генов на N- или С-конце, мы создали серию минималистичных остовов векторов-таргетингов, обеспечивающих плавную одноэтапную интеграцию обоих плеч гомологии с использованием сборки Golden Gate на основе SapI [4]. (рис. 2А). Направленные векторные скелеты, несущие современные мономерные флуоресцентные белки, доступны от Addgene, включая необязательные P2A-связанные элементы селекции (рис. 2B). С помощью наших оптимизированных векторных остовов целевые векторы могут быть собраны за то же время клонирования, которое требуется для вставки олигонуклеотидов гРНК в экспрессионные векторы Cas9.Следовательно, при использовании наших векторных остовов для ITPN аллели флуоресцентных репортеров в клеточных линиях и моделях органоидов могут быть созданы всего за 2 недели, включая процедуры молекулярного клонирования для сборки вектора (рис. 2C). Мы обобщили наши рекомендации по дизайну нок-ин и одноэтапной сборке вектора таргетинга в протоколе (файл S1). Кроме того, за короткий промежуток времени могут быть созданы новые варианты нацеливания на векторные остовы, например, замена донора другой последовательностью флуоресцентного белка (файл S2).
(рис. 2А). Направленные векторные скелеты, несущие современные мономерные флуоресцентные белки, доступны от Addgene, включая необязательные P2A-связанные элементы селекции (рис. 2B). С помощью наших оптимизированных векторных остовов целевые векторы могут быть собраны за то же время клонирования, которое требуется для вставки олигонуклеотидов гРНК в экспрессионные векторы Cas9.Следовательно, при использовании наших векторных остовов для ITPN аллели флуоресцентных репортеров в клеточных линиях и моделях органоидов могут быть созданы всего за 2 недели, включая процедуры молекулярного клонирования для сборки вектора (рис. 2C). Мы обобщили наши рекомендации по дизайну нок-ин и одноэтапной сборке вектора таргетинга в протоколе (файл S1). Кроме того, за короткий промежуток времени могут быть созданы новые варианты нацеливания на векторные остовы, например, замена донора другой последовательностью флуоресцентного белка (файл S2).
Рис. 2. Одноэтапная сборка нацеливающего вектора и ITPN ускоряют мечение флуоресцентных генов.
(A) Схематический план одноэтапной сборки ТВ с помощью лигирования гомологических плеч на основе SapI, опосредованного золотыми воротами. Плечи гомологии могут быть амплифицированы из геномной ДНК или упорядочены в виде коммерчески синтезированных фрагментов ДНК. (B) Обзор каркасных конструкций, доступных в Addgene. Вставные скелеты содержат один из 4 различных флуоресцентных белков и необязательные кассеты резистентности, связанные с P2A.Как указано, остовные конструкции подходят для нокаута либо на С-, либо на N-конце. (C) Схематический рабочий процесс с изложением мечения флуоресцентных генов в органоидах с использованием ITPN. После электропорации органоидам обычно требуется примерно 10 дней роста перед FACS-очисткой флуоресцентных нокаутных клеток. В качестве альтернативы, флуоресцентные клональные органоиды могут быть собраны вручную и объединены. Проверка последовательности индивидуально подобранных клональных линий не требуется при редактировании через ITPN. ITPN, парный никинг в трансе; ТВ, вектор наведения.
https://doi.org/10.1371/journal.pbio.3001527.g002
Чтобы проверить эффективность ITPN с использованием наших недавно разработанных векторов нацеливания, мы создали тройные флуоресцентные нокауты путем одновременного нацеливания на 3 отдельных геномных локуса. В частности, мы нацелили С-конец локуса HIST1h3BC на нокаут mTurquoise2-P2A-puromycin R , С-конец локуса CDh2 на нокаут mScarlet, и в качестве третьего локуса мы включал N-концевой нокаут mNeongreen либо в локусе LMNA , SEC61B , либо в локусе MAP4 (фиг. 3A).Коктейли ДНК, содержащие различные комбинации нацеливающих векторов и соответствующих им экспрессионных конструкций Cas9, подвергали электропорации во фракционированные опухолевые органоиды. Органоидам давали возможность сформироваться в течение 10 дней без отбора пуромицином перед количественной оценкой эффективности «сырого нокаута» с помощью проточного анализа отдельных клеток. Как и ожидалось, во всех трех условиях во фракциях нокаунов преобладали клетки, несущие одиночные нокауты в любом из целевых генов. Однако мы легко обнаружили клетки, несущие множественные нокауты, включая клетки, в которых все 3 гена редактировались одновременно (рис. 3В).Общая эффективность нокаута для каждого гена-мишени и доля клеток, несущих множественные нокауты, суммированы на рис. 3С. Чтобы подтвердить достоверность слияния генов, мы создали поликлональные линии с тройным нокаутом из каждого условия редактирования путем ручного выбора и объединения клональных тройных положительных органоидов. Анализ TIDE снова подтвердил отсутствие целевых вставок в нецелевых аллелях всех отредактированных генов (S5 Fig). Кроме того, мы подтвердили предполагаемую интеграцию нокаута с помощью секвенирования по Сэнгеру (рис. S6).Затем мы зафиксировали ночной рост нашей линии нокаута TKI-3, используя визуализацию живых клеток, чтобы продемонстрировать нормальное поведение роста и фенотип (рис. 3D). Каждый канал может быть записан без чрезмерного обесцвечивания, что позволяет получить многомерное динамическое считывание хромосомной нестабильности во время митоза, включая ошибки хроматина (h3B1C), сборку веретена (MAP4) и дефекты мембраны или бинуклеацию (CDh2).
Однако мы легко обнаружили клетки, несущие множественные нокауты, включая клетки, в которых все 3 гена редактировались одновременно (рис. 3В).Общая эффективность нокаута для каждого гена-мишени и доля клеток, несущих множественные нокауты, суммированы на рис. 3С. Чтобы подтвердить достоверность слияния генов, мы создали поликлональные линии с тройным нокаутом из каждого условия редактирования путем ручного выбора и объединения клональных тройных положительных органоидов. Анализ TIDE снова подтвердил отсутствие целевых вставок в нецелевых аллелях всех отредактированных генов (S5 Fig). Кроме того, мы подтвердили предполагаемую интеграцию нокаута с помощью секвенирования по Сэнгеру (рис. S6).Затем мы зафиксировали ночной рост нашей линии нокаута TKI-3, используя визуализацию живых клеток, чтобы продемонстрировать нормальное поведение роста и фенотип (рис. 3D). Каждый канал может быть записан без чрезмерного обесцвечивания, что позволяет получить многомерное динамическое считывание хромосомной нестабильности во время митоза, включая ошибки хроматина (h3B1C), сборку веретена (MAP4) и дефекты мембраны или бинуклеацию (CDh2).
Рис. 3. Мультиплексное флуоресцентное мечение генов в органоидах человека с использованием ITPN.
(A) Мультиплексное мечение флуоресцентных генов в опухолевых органоидах толстой кишки человека в 3 различных геномных локусах с использованием ITPN.С-концевая интеграция mScarlet в локусе CDh2 и mTurquoise2-P2A-Puromycin в локусе HIST1h3BC была объединена с N-концевой интеграцией mNeongreen либо в LMNA , SEC61B , либо в MAP . На схемах: позиции никеля Cas9 D10A (красные треугольники) и мотивы, прилегающие к протоспейсеру (черные столбцы), указаны для каждой нокаутной конструкции, а также используемая гРНК (зеленая стрелка). Органоиды подвергали электропорации одновременно со всеми 3 векторами-мишенями для получения одностадийных мультиплексированных тройных нокинов.(B) Все 3 комбинации нацеливания дали тройные популяции нокаута с практической эффективностью, как показал анализ потока (числа указывают частоту (%) нокаутированных клеток во всей целевой клеточной популяции).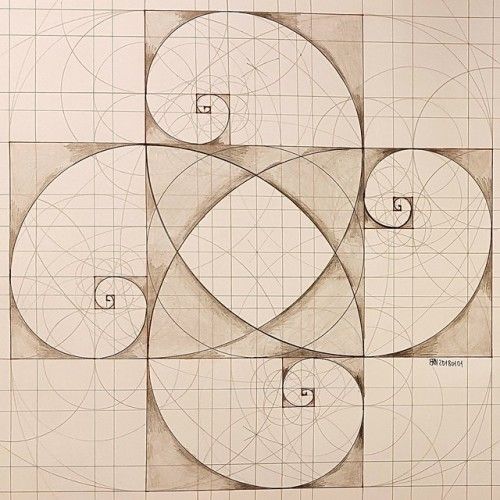 Снимки изображений показывают ожидаемую субклеточную локализацию каждого слитого белка (масштабная линейка = 10 мкм). Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ). (C) Обзор эффективности мечения мультиплексированных генов, определяемой анализом проточной цитометрии. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ).(D) Визуализация живых клеток опухолевых органоидов толстой кишки человека, несущих нокауты CDh2-mScarlet , HIST1h3BC-mTurquoise2 и mNeongreen-MAP4 . На верхней панели показаны репрезентативные кадры роста органоидов с течением времени (масштабная линейка = 10 мкм). Для отделов I и II снимки каждого канала показаны в метафазе и анафазе (масштабная линейка = 5 мкм). ITPN, парный никинг в трансе.
Снимки изображений показывают ожидаемую субклеточную локализацию каждого слитого белка (масштабная линейка = 10 мкм). Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ). (C) Обзор эффективности мечения мультиплексированных генов, определяемой анализом проточной цитометрии. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ).(D) Визуализация живых клеток опухолевых органоидов толстой кишки человека, несущих нокауты CDh2-mScarlet , HIST1h3BC-mTurquoise2 и mNeongreen-MAP4 . На верхней панели показаны репрезентативные кадры роста органоидов с течением времени (масштабная линейка = 10 мкм). Для отделов I и II снимки каждого канала показаны в метафазе и анафазе (масштабная линейка = 5 мкм). ITPN, парный никинг в трансе.
https://doi.org/10.1371/journal.pbio.3001527.g003
В совокупности эти результаты показывают, что ITPN поддерживает высокий уровень точности для различных геномных локусов и позволяет мультиплексировать флуоресцентную маркировку генов в органоидах человека. Используя обычные протоколы редактирования, создание линий органоидов, несущих несколько флуоресцентных нокаутов, очень трудоемко. Используя ITPN, органоиды с несколькими правками могут быть созданы в течение 2 недель. В качестве альтернативы, в случае неудачной попытки мультиплексного нацеливания генов, клетки с одним нокаутом могут быть объединены и перенацелены. Более того, мы создали те же комбинации тройных нокаутов в 2 раундах нацеливания и использовали промежуточную селекцию антибиотиков для обогащения нокаутными клетками вместо ручного сбора (S7 Fig).
Используя обычные протоколы редактирования, создание линий органоидов, несущих несколько флуоресцентных нокаутов, очень трудоемко. Используя ITPN, органоиды с несколькими правками могут быть созданы в течение 2 недель. В качестве альтернативы, в случае неудачной попытки мультиплексного нацеливания генов, клетки с одним нокаутом могут быть объединены и перенацелены. Более того, мы создали те же комбинации тройных нокаутов в 2 раундах нацеливания и использовали промежуточную селекцию антибиотиков для обогащения нокаутными клетками вместо ручного сбора (S7 Fig).
Поскольку целостность последовательности нецелевого аллеля, который не несет нокаут, сохраняется при редактировании через ITPN, этот вторичный аллель может быть перенацелен с использованием того же локус-специфического нацеливающего вектора для получения гомозиготных нокаутов. Это также обеспечивает прямую дифференциальную модификацию материнских и отцовских аллелей, предлагая 2 разных вектора нацеливания для одного и того же локуса. Чтобы выяснить, допускает ли ITPN одновременное образование биаллельных нокинов, несущих разные флуоресцентные метки в каждом аллеле, мы нацелились на локусы SEC61B , MAP4 и HIST1h3BC в опухолевых органоидах с векторами нацеливания mNeongreen и mScarlet.Анализ потока через 10 дней после электропорации подтвердил наличие двойной положительной клеточной популяции для каждого целевого локуса (рис. 4А). Генотипирование отобранных вручную линий подтвердило правильную модификацию каждой аллели (рис. S8). Кроме того, визуализация двуаллельных нокаутированных органоидов подтвердила обнаружение обоих аллель-специфических репортеров (рис. 4В). Затем мы выполнили визуализацию живых клеток органоидов и оценили биаллельный флуоресцентный выход (зеленый против красного) для отдельных клеток с течением времени (рис. 4C), как прямую демонстрацию того, как дифференциальные аллель-специфические модификации могут быть использованы для изучения аллель-специфической экспрессии.
Чтобы выяснить, допускает ли ITPN одновременное образование биаллельных нокинов, несущих разные флуоресцентные метки в каждом аллеле, мы нацелились на локусы SEC61B , MAP4 и HIST1h3BC в опухолевых органоидах с векторами нацеливания mNeongreen и mScarlet.Анализ потока через 10 дней после электропорации подтвердил наличие двойной положительной клеточной популяции для каждого целевого локуса (рис. 4А). Генотипирование отобранных вручную линий подтвердило правильную модификацию каждой аллели (рис. S8). Кроме того, визуализация двуаллельных нокаутированных органоидов подтвердила обнаружение обоих аллель-специфических репортеров (рис. 4В). Затем мы выполнили визуализацию живых клеток органоидов и оценили биаллельный флуоресцентный выход (зеленый против красного) для отдельных клеток с течением времени (рис. 4C), как прямую демонстрацию того, как дифференциальные аллель-специфические модификации могут быть использованы для изучения аллель-специфической экспрессии. уровней [18,19].Это доказательство принципа подчеркивает способность ITPN создавать аллель-специфические данные, которые, в зависимости от нокаут-матрицы, могут быть применены для решения многих биологических вопросов, начиная от аллель-специфических паттернов экспрессии и заканчивая дифференциальными биохимическими свойствами между диким типом и диким типом. мутантные белки.
уровней [18,19].Это доказательство принципа подчеркивает способность ITPN создавать аллель-специфические данные, которые, в зависимости от нокаут-матрицы, могут быть применены для решения многих биологических вопросов, начиная от аллель-специфических паттернов экспрессии и заканчивая дифференциальными биохимическими свойствами между диким типом и диким типом. мутантные белки.
Рис. 4. Одноэтапные дифференциальные флуоресцентные нокауты в одном локусе.
(A) Локусы SEC61B , MAP4 и HIST1h3BC модели опухолевого органоида, полученной от пациента, были нацелены через ITPN с векторами нацеливания mNeongreen и mScarlet в соответствии со схемой, показанной на рис. 3A.Анализ потока проводили через 10 дней после электропорации. Указан процент клеток, несущих одиночный нокаут, и клеток, несущих оба нокаута. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ). (B) Репрезентативные кадры опухолевых органоидов, несущих биаллельные модификации mNeongreen и mScarlet в локусах SEC61B , MAP4 и HIST1h3BC (масштабная линейка = 10 мкм). (C) Аллель-специфическое считывание в органоидах колоректального рака человека, содержащих дифференциально помеченные аллели HIST1h3BC .Органоиды, несущие нок-ины mNeongreen и mScarlet в локусе HIST1h3BC , визуализировали как живые клетки в течение 18 часов. На верхних панелях показаны кадры зелено-красной композиции с течением времени. На левом нижнем графике показаны изменения аллель-специфического выхода с течением времени для одной репрезентативной клетки. На график нанесены скорректированные флуоресцентные сигналы mScarlet, mNeongreen, а также отношение mScarlet/mNeongreen. Правый нижний график показывает 7 из 17 проанализированных соотношений одиночных клеток (mScarlet/mNeongreen).Базовые данные представлены в S1 Data. ITPN, парный никинг в трансе.
(C) Аллель-специфическое считывание в органоидах колоректального рака человека, содержащих дифференциально помеченные аллели HIST1h3BC .Органоиды, несущие нок-ины mNeongreen и mScarlet в локусе HIST1h3BC , визуализировали как живые клетки в течение 18 часов. На верхних панелях показаны кадры зелено-красной композиции с течением времени. На левом нижнем графике показаны изменения аллель-специфического выхода с течением времени для одной репрезентативной клетки. На график нанесены скорректированные флуоресцентные сигналы mScarlet, mNeongreen, а также отношение mScarlet/mNeongreen. Правый нижний график показывает 7 из 17 проанализированных соотношений одиночных клеток (mScarlet/mNeongreen).Базовые данные представлены в S1 Data. ITPN, парный никинг в трансе.
https://doi.org/10.1371/journal.pbio.3001527.g004
Обсуждение
Здесь мы показываем, что большие нок-ины, такие как флуоресцентные генные метки, могут быть созданы в органоидах человека с высокой эффективностью и точностью с использованием ITPN.![]() Эта стратегия превосходит обычное редактирование генома, опосредованное Cas9, поскольку ITPN не зависит от DSB, что сводит к минимуму риск нежелательных мутаций как в потенциальных нецелевых сайтах, так и в нецелевом аллеле, который не несет нокаут.Это также означает, что ITPN идеально подходит для введения гетерозиготных патогенных (зародышевых) мутаций. Дополнительным преимуществом является то, что проверка последовательности индивидуально отобранных клональных линий становится устаревшей, и, таким образом, все сгенерированные de novo клетки с нокаутом могут быть немедленно объединены. Это ускоряет распространение ранней культуры и сокращает общее время генерации нок-ин моделей. Кроме того, объединение всех созданных de novo нокаутированных клеток сохраняет генетическую гетерогенность, присутствующую в исходной культуре, что важно при работе с моделями опухолевых органоидов, полученных от пациентов.Чтобы обойти трудоемкое молекулярное клонирование векторов-мишеней, мы создали палитру векторных остовов, которые можно настроить по локусу за то же время клонирования, которое требуется для вставки олиго гРНК в экспрессионные векторы Cas9.
Эта стратегия превосходит обычное редактирование генома, опосредованное Cas9, поскольку ITPN не зависит от DSB, что сводит к минимуму риск нежелательных мутаций как в потенциальных нецелевых сайтах, так и в нецелевом аллеле, который не несет нокаут.Это также означает, что ITPN идеально подходит для введения гетерозиготных патогенных (зародышевых) мутаций. Дополнительным преимуществом является то, что проверка последовательности индивидуально отобранных клональных линий становится устаревшей, и, таким образом, все сгенерированные de novo клетки с нокаутом могут быть немедленно объединены. Это ускоряет распространение ранней культуры и сокращает общее время генерации нок-ин моделей. Кроме того, объединение всех созданных de novo нокаутированных клеток сохраняет генетическую гетерогенность, присутствующую в исходной культуре, что важно при работе с моделями опухолевых органоидов, полученных от пациентов.Чтобы обойти трудоемкое молекулярное клонирование векторов-мишеней, мы создали палитру векторных остовов, которые можно настроить по локусу за то же время клонирования, которое требуется для вставки олиго гРНК в экспрессионные векторы Cas9. Кроме того, наш дизайн основы вектора таргетинга является модульным, так что сам шаблон донора можно легко адаптировать к более сложным и изощренным конструкциям репортеров [5]. Наши векторные магистрали доступны на Addgene, а наши протоколы (файлы S1 и S2) содержат подробные инструкции по их применению.
Кроме того, наш дизайн основы вектора таргетинга является модульным, так что сам шаблон донора можно легко адаптировать к более сложным и изощренным конструкциям репортеров [5]. Наши векторные магистрали доступны на Addgene, а наши протоколы (файлы S1 и S2) содержат подробные инструкции по их применению.
Хотя абсолютная эффективность нок-ина зависит от многих переменных, наиболее практичным фактором, определяющим успешную стратегию, является то, что нок-ин постоянно получается с каждой попыткой. Чтобы продемонстрировать надежную эффективность ITPN в органоидах, мы создали тройные нокины в 3 независимых геномных локусах в одном раунде таргетинга и выполнили одноэтапное создание биаллельных нокинов, несущих аллель-специфические репортеры. В наших руках эффективность ITPN превосходит стратегии CRISPR, в которых используются обычные шаблоны доноров.В соответствии с более ранними отчетами [8, 9, 14, 15], важной переменной, влияющей на общую эффективность, был ITPN или ITPC донора. Хотя нокауты, опосредованные NHEJ, с помощью таких стратегий, как CRISPR-HOT [6], сводят к минимуму потребность в молекулярном клонировании, наши результаты показывают, что ITPN значительно повышает эффективность и точность по сравнению со стратегиями, опосредованными NHEJ, в то время как наши выделенные векторные основы сводят к минимуму молекулярное клонирование. в аналогичной степени.
в аналогичной степени.
Из-за почти точного сходства последовательностей материнских и отцовских аллелей различия в характере экспрессии между двумя аллелями данного гена плохо изучены.Точно так же изменения внутриклеточной локализации, белковых взаимодействий и/или биохимических свойств между белками дикого типа и мутантными белками, такими как онкогены, редко исследуются в одних и тех же клетках из-за практически невыполнимой задачи модификации обоих аллелей независимо друг от друга разными метками. Поскольку ITPN генерирует нокаут-аллели без модификации нецелевого аллеля, тот же локус может быть перенацелен с помощью альтернативного донорского шаблона. В качестве доказательства принципа мы создали биаллельный нокаут с различными флуорофорами в локусах HIST1h3BC , SEC61B и MAP4 в одном раунде нацеливания.Точная оценка аллельного дисбаланса важна для понимания генетических и эпигенетических механизмов регуляции генов и нарушения регуляции во время канцерогенеза.
В заключение, ITPN представляет собой универсальную стратегию, которая позволяет быстро и эффективно создавать нокауты в органоидах человека. Мы предполагаем, что наш подход может быть легко применен к моделям органоидов, полученных из других тканей или источников, таких как модели органоидов, полученные из плюрипотентных стволовых клеток. На сегодняшний день сообщалось о различных CRISPR-опосредованных стратегиях включения, которые достигают достаточной эффективности, чтобы сделать генетическое редактирование практичным в моделях органоидов.ITPN сопоставим с точки зрения эффективности, но выделяется тем, что не зависит от DSB и, следовательно, обладает высочайшей внутренней точностью точного редактирования генома. В сочетании с бесшовной одноэтапной генерацией векторов наведения ITPN представляет собой важный технологический прогресс в создании высокоточных систем моделирования.
Материалы и методы
Вектор в сборе
Скелеты вектора-мишени были созданы с помощью бесшовной сборки на основе рекомбиназы (клонирование In-Fusion, Takara Biotech) из коммерчески синтезированного фрагмента ДНК (IDT или Genscript), несущего mNeongreen [20], mTurquiose2 [21], mScarlet [22] или фотоконвертируемый Dendra2 [23] с необязательными элементами P2A-селекции (таблица S2) в ПЦР-амплифицированный общий фрагмент основной цепи (FWD: tcctcgctcactgactcgct, REV: gcggtattttctccttacgcatctg). См. Файл S2 для подробного объяснения того, как создавать новые опорные точки вектора таргетинга. Чтобы создать локус-специфические нацеливающие векторы, коммерчески синтезированные гомологические фрагменты плеча (таблица S3) были вставлены в нацеливающие векторные скелеты с использованием сборки золотых ворот на основе SapI, как описано ранее [4]; см. файл S1 для более подробного протокола. Cas9 дикого типа (addgene #48139) и Cas9 D10A nickase (addgene #48141) локус-специфические векторы экспрессии были созданы в соответствии с опубликованными протоколами [24].
См. Файл S2 для подробного объяснения того, как создавать новые опорные точки вектора таргетинга. Чтобы создать локус-специфические нацеливающие векторы, коммерчески синтезированные гомологические фрагменты плеча (таблица S3) были вставлены в нацеливающие векторные скелеты с использованием сборки золотых ворот на основе SapI, как описано ранее [4]; см. файл S1 для более подробного протокола. Cas9 дикого типа (addgene #48139) и Cas9 D10A nickase (addgene #48141) локус-специфические векторы экспрессии были созданы в соответствии с опубликованными протоколами [24].
Органоидная культура
Опухолевой органоид пациента с идентификатором P9T (PDTO-9) был получен из ранее опубликованного биобанка колоректального рака [16]. PDTO-9 поддерживали при 37°C в атмосфере 5% CO 2 с затравкой экстрактом базальной мембраны RGF (BME), тип 2 (Cultrex). Культуральные среды состояли из улучшенной среды DMEM/F12 (Gibco) с добавлением пенициллина-стрептомицина (Lonza, 10 ед. мл -1 ), GlutaMAX (Gibco, 1x), буфера HEPES (Gibco, 10 мМ), среды, кондиционированной Noggin (10 мМ). %), кондиционированная R-spondin1 среда (10%), B-27 (Gibco, 1x), никотинамид (Sigma-Aldrich, 10 мМ), N -ацетилцистеин (Sigma-Aldrich, 1.25 мМ), SB202190 (Gentaur, 10 мкМ), A83-01 (Tocris, 500 нМ) и рекомбинантный EGF человека (PeproTech, 50 нг мл -1 ). Культуры PDTO-9 пассировали еженедельно и поддерживали ниже пассажа 10. Вкратце, PDTO диссоциировали с использованием трипсин-ЭДТА (Sigma-Aldrich) и высевали в BME в предварительно нагретый 24-луночный планшет. Ингибитор ROCK Y-27632 (Gentaur, 10 мкМ) добавляли в культуральную среду после посева на 2 дня.
%), кондиционированная R-spondin1 среда (10%), B-27 (Gibco, 1x), никотинамид (Sigma-Aldrich, 10 мМ), N -ацетилцистеин (Sigma-Aldrich, 1.25 мМ), SB202190 (Gentaur, 10 мкМ), A83-01 (Tocris, 500 нМ) и рекомбинантный EGF человека (PeproTech, 50 нг мл -1 ). Культуры PDTO-9 пассировали еженедельно и поддерживали ниже пассажа 10. Вкратце, PDTO диссоциировали с использованием трипсин-ЭДТА (Sigma-Aldrich) и высевали в BME в предварительно нагретый 24-луночный планшет. Ингибитор ROCK Y-27632 (Gentaur, 10 мкМ) добавляли в культуральную среду после посева на 2 дня.
Нормальные органоиды толстой кишки человека выдерживали при 37°C в атмосфере с 5% CO 2 и засевали матригелем с пониженным содержанием фактора роста (BD Biosciences).Питательные среды [25] состояли из улучшенной среды DMEM/F12 (Gibco) с добавлением пенициллина-стрептомицина (Lonza, 10 ед. мл -1 ), GlutaMAX (Gibco, 1x), буфера HEPES (Gibco, 10 мМ), кондиционированного Noggin. среда (10%), кондиционированная R-spondin1 среда (20%), B-27 (Gibco, 1x), N -ацетилцистеин (Sigma-Aldrich, 1,25 мМ), A83-01 (Tocris, 500 нМ), рекомбинантный EGF человека (PeproTech, 50 нг мл -1 ), рекомбинантный IGF-1 человека (BioLegend, 100 нг/мл), рекомбинантный основной FGF человека (Peprotech, FGF-2 50 нг/мл) и 0. 5 нМ суррогата Wnt (экспресс U-белка). Органоиды пассировали, как описано выше.
5 нМ суррогата Wnt (экспресс U-белка). Органоиды пассировали, как описано выше.
Электропорация органоидов
Для создания нок-инов в нормальных или колоректальных опухолевых органоидах 1 × 10 6 клеток примерно из 5 клеточных скоплений подвергали коэлектропорации с 15 мкг ДНК в соотношении 1:1 Cas9 (Addgene #48139) или никазы Cas9 D10A. (Addgene # 48141) и вектор-мишень с использованием суперэлектропоратора NEPA21 (Nepagene) в соответствии с описанными условиями [26].
Анализ потока
Органоидыобрабатывали трипсином и фильтровали через сито CellTrics 10 мкм (Sysmex) для получения суспензии одиночных клеток.Для количественной оценки эффективности нокаута клетки подвергали анализу потока (FACSCelesta, BD) не менее чем через 10 дней после электропорации. Ворота были установлены на основе отрицательного контроля/популяции.
Генотипирование и анализ TIDE
Поликлональные или клональные нокаут-культуры были созданы путем ручного сбора или FACS. Сайт-специфические интеграции были подтверждены с помощью ПЦР генотипирования экстракта геномной ДНК с использованием наборов локус-специфических праймеров (таблица S4) с последующим секвенированием по Сэнгеру. Анализ TIDE выполняли на основе данных секвенирования по Сэнгеру вторичных «не нокаутированных» аллелей с использованием считывания секвенирования по Сэнгеру линии родительских органоидов в качестве хроматограммы контрольного образца.
Сайт-специфические интеграции были подтверждены с помощью ПЦР генотипирования экстракта геномной ДНК с использованием наборов локус-специфических праймеров (таблица S4) с последующим секвенированием по Сэнгеру. Анализ TIDE выполняли на основе данных секвенирования по Сэнгеру вторичных «не нокаутированных» аллелей с использованием считывания секвенирования по Сэнгеру линии родительских органоидов в качестве хроматограммы контрольного образца.
Полногеномное секвенирование и картирование прочтений
Геномную ДНК 3 клонально расширенных линий органоидов SEC61B -mScarlet, опосредованных ITPN, выделяли с использованием набора QIAamp DNA Micro Kit в соответствии с инструкциями производителя. Библиотеки секвенирования Illumina были созданы с использованием 200 нг геномной ДНК в соответствии со стандартными протоколами (Illumina). После WGS до базового охвата 15x (Illumina NovaSeq 6000, 2 × 150 п.н.) была выполнена первоначальная обработка считываний последовательностей с использованием полного конвейера анализа, доступного на https://github. com/UMCUGenetics/NF-IAP. Вкратце, инструмент картирования Burrows-Wheeler Aligner v0.7.17 использовался для сопоставления считываний последовательностей с эталонным геномом человека GRCh48 с настройками «bwa mem -c 100 –M» [27]. Затем дубликаты чтения были помечены с помощью Sambamba v0.6.8, а для повторного выравнивания использовался набор инструментов для анализа генома (GATK) v4.1.3.0 [28].
com/UMCUGenetics/NF-IAP. Вкратце, инструмент картирования Burrows-Wheeler Aligner v0.7.17 использовался для сопоставления считываний последовательностей с эталонным геномом человека GRCh48 с настройками «bwa mem -c 100 –M» [27]. Затем дубликаты чтения были помечены с помощью Sambamba v0.6.8, а для повторного выравнивания использовался набор инструментов для анализа генома (GATK) v4.1.3.0 [28].
Вызов вариантов и фильтрация
Затем были вызваны несколько вариантов с помощью GATK HaplotypeCaller v4.1.3.0 и GATK-Queue v.4.1.3.0 на основе настроек по умолчанию и дополнительной опции «EMIT_ALL_CONFIDENT_SITES.Впоследствии для оценки качества позиций вариантов использовался GATK VariantFiltration v4.1.3.0 с параметрами -snpFilterName SNP_LowQualityDepth -snpFilterExpression «QD < 2.0" -snpFilterName SNP_MappingQuality -snpFilterExpression "MQ < 40.0" -snpFilterName SNP_StrandBias > 60.0» -snpFilterName SNP_HaplotypeScoreHigh -snpFilterExpression «HaplotypeScore > 13.0» -snpFilterName SNP_MQRankSumLow -snpFilterExpression «MQRankSum < −12. 5» -snpFilterName SNP_ReadPosRankSumLow -snpFilterExpression «ReadPosRankSum»0» -snpFilterName SNP_HardToValidate -snpFilterExpression «MQ0 > = 4 && ((MQ0 / (1.0 * DP)) > 0.1)» -snpFilterName SNP_LowCoverage -snpFilterExpression «DP < 5» -snpFilterName SNP_VeryLowQual -snpFilterQualName SNP0Lows «QUAL» <3 -snpFilterExpression «QUAL > = 30,0 && QUAL < 50,0» -snpFilterName SNP_SOR -snpFilterExpression «SOR > 4,0» -cluster 3 -window 10 -indelType INDEL -indelType MIXED -indelFilterName INDEL_LowQualityDepth -indelFilterExpression «QD < 2.0» -indelFilterName INDEL_StrandBias -indelFilterExpression «FS > 200.0» -indelFilterExpression INDEL_ReadPosRankSumLow -indelFilterExpression «ReadPosRankSum < −20.0» -indelFilterName INDEL_HardToValidate -indelFilterExpression «MQ0 > = 4 &DP& ((MQ0 *0 / (1.1.) INDEL_LowCoverage -indelFilterExpression «DP < 5» -indelFilterName INDEL_VeryLowQual -indelFilterExpression «QUAL < 30,0» -indelFilterName INDEL_LowQual -indelFilterExpression «QUAL > = 30,0 && QUAL < 50.
5» -snpFilterName SNP_ReadPosRankSumLow -snpFilterExpression «ReadPosRankSum»0» -snpFilterName SNP_HardToValidate -snpFilterExpression «MQ0 > = 4 && ((MQ0 / (1.0 * DP)) > 0.1)» -snpFilterName SNP_LowCoverage -snpFilterExpression «DP < 5» -snpFilterName SNP_VeryLowQual -snpFilterQualName SNP0Lows «QUAL» <3 -snpFilterExpression «QUAL > = 30,0 && QUAL < 50,0» -snpFilterName SNP_SOR -snpFilterExpression «SOR > 4,0» -cluster 3 -window 10 -indelType INDEL -indelType MIXED -indelFilterName INDEL_LowQualityDepth -indelFilterExpression «QD < 2.0» -indelFilterName INDEL_StrandBias -indelFilterExpression «FS > 200.0» -indelFilterExpression INDEL_ReadPosRankSumLow -indelFilterExpression «ReadPosRankSum < −20.0» -indelFilterName INDEL_HardToValidate -indelFilterExpression «MQ0 > = 4 &DP& ((MQ0 *0 / (1.1.) INDEL_LowCoverage -indelFilterExpression «DP < 5» -indelFilterName INDEL_VeryLowQual -indelFilterExpression «QUAL < 30,0» -indelFilterName INDEL_LowQual -indelFilterExpression «QUAL > = 30,0 && QUAL < 50. 0” -indelFilterName INDEL_SOR -indelFilterExpression “SOR > 10.0.”
0” -indelFilterName INDEL_SOR -indelFilterExpression “SOR > 10.0.”
Мутации низкого качества и субклональные мутации, накопленные в ходе клональной экспансии органоидных линий, были исключены путем аннотирования с помощью SMuRF версии 2.1.2, как описано [29] (https://github.com/ToolsVanBox/SMuRF). Мы включили все варианты в каждый клон с аутосомными или Х-хромосомами, присутствующими менее чем в 3 клональных образцах, прошедших VariantFiltration, с показателем качества по шкале GATK phred ≥60; минимальное базовое покрытие 5X, качество картирования ≥30 и частота вариантного аллеля не менее 0.15 [29,30]. Вызов структурных вариаций выполнялся с помощью конвейера GRIDSS-purple-linx версии 1.3.2 с использованием всех парных комбинаций 3 образцов WGS в качестве пар опухоль-нормальность [31].
Анализ предсказанных in silico нецелевых областей
Все потенциальные нецелевые области протоспейсера для направляющей последовательности 5′-GGGGTCGGACCAGGCTGTAG-3′ были предсказаны с использованием общедоступного инструмента CasOFFinder [32], с использованием NGG PAM и с учетом до 4 несоответствий. В качестве областей интереса рассматривались как потенциальные нецелевые области протоспейсера, так и фланкирующие 200 оснований.С помощью BEDtools v2.27.1 все варианты, прошедшие фильтрацию SMuRF, пересекались с интересующими областями [33]. Кроме того, все начальные и конечные координаты структурных вариаций, названных GRIDSS-purple-linx, пересекались с одними и теми же потенциальными нецелевыми областями генома.
В качестве областей интереса рассматривались как потенциальные нецелевые области протоспейсера, так и фланкирующие 200 оснований.С помощью BEDtools v2.27.1 все варианты, прошедшие фильтрацию SMuRF, пересекались с интересующими областями [33]. Кроме того, все начальные и конечные координаты структурных вариаций, названных GRIDSS-purple-linx, пересекались с одними и теми же потенциальными нецелевыми областями генома.
Визуализация живых органоидов
Для поддержки микроскопии живых клеток органоидов PDTO-9 или нормальные культуры органоидов пассировали за 5–7 дней до визуализации. PDTO собирали за 24 часа до визуализации и ресуспендировали в охлажденной льдом смеси культуральных сред, содержащих 50% об./об. BME или 90% об./об. Matrigel.Суспензию органоидов затем высевали в чашку со льдом со стеклянным дном WillCo (WillCo Wells B.V.), покрытую тонкой пленкой BME или Matrigel. Органоидам давали осесть на льду перед полимеризацией геля при 37°C и добавлением культуральной среды.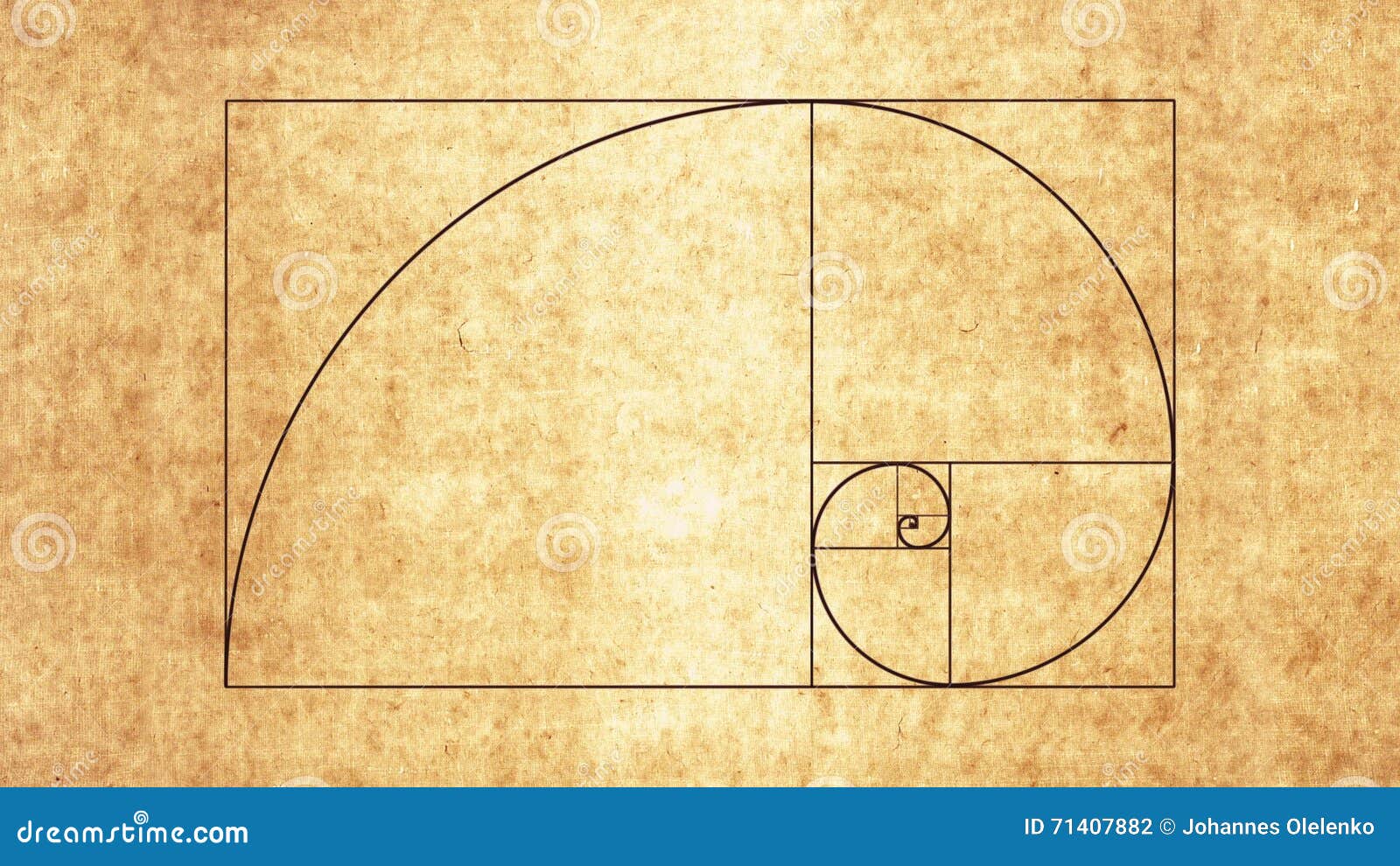 Нарост снимали в течение ночи с помощью конфокальной системы с вращающимся диском (Nikon, частота кадров 15 минут, шаг по оси z 1,4 мкм). Данные визуализации были проанализированы с помощью Fiji (ImageJ). Для биаллельных органоидов HIST1h3BC был использован специальный макрос анализа [34] для отслеживания отдельных клеток и мониторинга их сигналов mNeongreen и mScarlet.Коррекция отбеливания (на канал, на момент времени) выполнялась на основе интегральных сигналов флуоресценции соответствующих органоидов.
Нарост снимали в течение ночи с помощью конфокальной системы с вращающимся диском (Nikon, частота кадров 15 минут, шаг по оси z 1,4 мкм). Данные визуализации были проанализированы с помощью Fiji (ImageJ). Для биаллельных органоидов HIST1h3BC был использован специальный макрос анализа [34] для отслеживания отдельных клеток и мониторинга их сигналов mNeongreen и mScarlet.Коррекция отбеливания (на канал, на момент времени) выполнялась на основе интегральных сигналов флуоресценции соответствующих органоидов.
Вспомогательная информация
S1 Рис. Анализы методом проточной цитометрии для измерения эффективности нокаута.
Показаны репрезентативные графики проточной цитометрии для анализа эффективности нокаута, включая неотредактированные контроли. Панели (A), (B), (C) и (D) относятся к рис. 1C, 1D, 1F и 1G соответственно. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ).ITPC, внутритранс-парное расщепление; ITPN, парный никинг в трансе; MMEJ, соединение концов, опосредованное микрогомологией; NHEJ, негомологичное соединение концов; вес, дикий тип.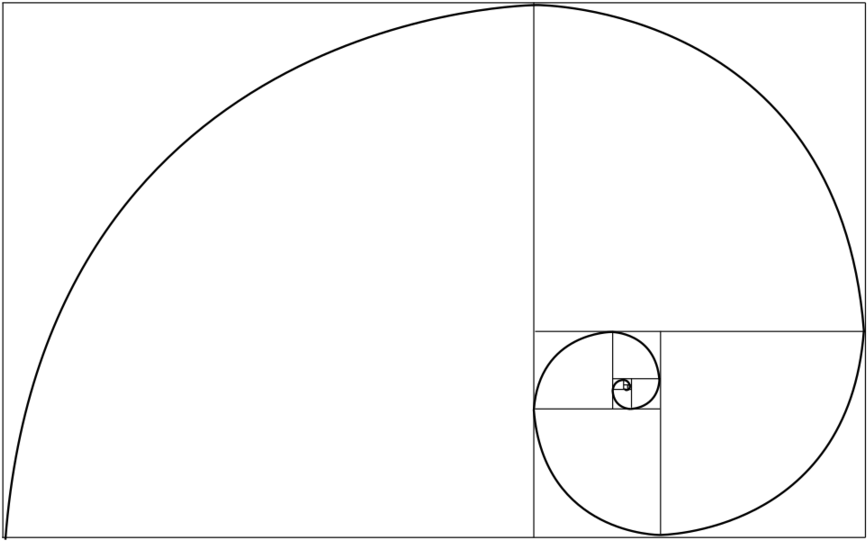
10.1371/journal.pbio.3001527.s001
(EPS)
S2 Рис. Целевой анализ TIDE различных стратегий нокаута в локусе
SEC61B . Для проведения анализа TIDE геномную ДНК экстрагировали из поликлональных популяций с нокаой, созданных с помощью MMEJ, традиционного нацеливания, ITPC (с Cas9 дикого типа) или ITPN (с никазой Cas9 D10A) (см. рис. 1C).Поликлональные линии были созданы путем ручного объединения приблизительно 10 клональных нокаутированных органоидов. Анализ TIDE выполняли с использованием считывания секвенирования по Сэнгеру модели опухолевого органоида, полученной от родительского пациента, в качестве хроматограммы контрольного образца. Состояние NHEJ, совместно перенесенное с Cas9 дикого типа, было исключено из этого анализа, так как было слишком мало доступных органоидов для получения поликлональной линии. Процент чтений, содержащих вставки, отображается в левом верхнем углу. Базовые данные представлены в S2 Data.ITPC, внутритранс-парное расщепление; ITPN, парный никинг в трансе; MMEJ, соединение концов, опосредованное микрогомологией; NHEJ, негомологичное соединение концов; WT, дикий тип.
10.1371/journal.pbio.3001527.s002
(EPS)
S3 Рис. Точность опосредованных ITPN флуоресцентных нокаутов в локусе
SEC61B .Секвенирование по Сэнгеру было выполнено на локусе SEC61B 11 клональных опухолевых органоидных нокаутных линий, полученных от пациентов, полученных с помощью ITPN (см. рис. 1C). Линии нокаута были созданы путем ручного отбора отдельных крупных клональных органоидов с нокаутом.Для каждого клона показаны 5′- и 3′-соединения аллеля нокаута и область-мишень «нецелевого» аллеля. Клон № 7 несет гомозиготный нок-ин mScarlet в локусе SEC61B и, следовательно, не содержит нецелевого аллеля. Базовые данные представлены в S2 Data. ITPN, парный никинг в трансе.
10.1371/journal.pbio.3001527.s003
(TIF)
S4 Рис. Нокаут KRT20-mScarlet в нормальных органоидах толстой кишки человека.
(A) Схематическое изображение стратегии нацеливания KRT20 на подавление mScarlet-P2A-Blasticidin R . Донор был фланкирован гомологическими плечами, соответствующими восходящим и нисходящим последовательностям С-конца локуса KRT20 человека. Никазу Cas9 D10A нацеливали вблизи стоп-кодона с использованием гРНК, как указано (зеленая стрелка). Показаны сайты расщепления никазы Cas9 D10A (треугольники) и мотивы, прилегающие к протоспейсеру (черная полоса). Гомология вверх и вниз по течению представлена синим и красным цветом соответственно. Вставные органоиды были получены путем электропорации нормальных скоплений органоидов толстой кишки человека с последующим ростом в течение 10 дней.Для отбора успешно нацеленных клеток органоиды обрабатывали в течение 7 дней бластицидином, после чего вручную собирали клональный органоид, содержащий красные флуоресцентные клетки. (B) Флуоресцентные изображения репрезентативных органоидов KRT20 -mScarlet-P2A-Blasticidin R , содержащих как KRT20-положительные, так и KRT20-отрицательные клетки (красный: KRT20-mScarlet). Масштабная линейка = 50 мкм.
Донор был фланкирован гомологическими плечами, соответствующими восходящим и нисходящим последовательностям С-конца локуса KRT20 человека. Никазу Cas9 D10A нацеливали вблизи стоп-кодона с использованием гРНК, как указано (зеленая стрелка). Показаны сайты расщепления никазы Cas9 D10A (треугольники) и мотивы, прилегающие к протоспейсеру (черная полоса). Гомология вверх и вниз по течению представлена синим и красным цветом соответственно. Вставные органоиды были получены путем электропорации нормальных скоплений органоидов толстой кишки человека с последующим ростом в течение 10 дней.Для отбора успешно нацеленных клеток органоиды обрабатывали в течение 7 дней бластицидином, после чего вручную собирали клональный органоид, содержащий красные флуоресцентные клетки. (B) Флуоресцентные изображения репрезентативных органоидов KRT20 -mScarlet-P2A-Blasticidin R , содержащих как KRT20-положительные, так и KRT20-отрицательные клетки (красный: KRT20-mScarlet). Масштабная линейка = 50 мкм. (C) Секвенирование по Сэнгеру проводили на линии нокаута KRT20 -mScarlet-P2A-Blasticidin R .5′- и 3′-соединения аллеля нокаута не повреждены. Базовые данные представлены в S2 Data.
(C) Секвенирование по Сэнгеру проводили на линии нокаута KRT20 -mScarlet-P2A-Blasticidin R .5′- и 3′-соединения аллеля нокаута не повреждены. Базовые данные представлены в S2 Data.
10.1371/journal.pbio.3001527.s004
(EPS)
S5 Рис. TIDE-анализ мультиплексированных тройных нокаутных линий в опухолевых органоидах толстой кишки человека.
TIDE-анализ «нецелевых» аллелей для всех целевых генов в одноэтапных линиях с тройным нокаутом (см. рис. 3). TIDE выполняли с использованием считывания секвенирования по Сэнгеру органоидной модели родительской опухоли толстой кишки в качестве хроматограммы контрольного образца.Аллель SEC61B Triple KI-2 не анализировали, поскольку он не прошел контроль качества. Базовые данные представлены в S2 Data.
10.1371/journal.pbio.3001527.s005
(EPS)
S6 Рис. Секвенирование по Сэнгеру 5′- и 3′-нокаутных аллельных соединений в тройных опухолевых органоидах толстой кишки.
Секвенирование по Сэнгеру было проведено на поликлональных тройных опухолевых линиях толстой кишки, полученных путем ручного сбора тройных положительных клональных органоидов. Соединения 5′ и 3′ каждого аллеля нокаута показаны для всех 3 мультиплексных условий нокаута (см. рис. 3).Базовые данные представлены в S2 Data.
Соединения 5′ и 3′ каждого аллеля нокаута показаны для всех 3 мультиплексных условий нокаута (см. рис. 3).Базовые данные представлены в S2 Data.
10.1371/journal.pbio.3001527.s006
(TIF)
S7 Рис. Двухэтапное тройное нок-инирование в органоидах рака толстой кишки человека с использованием обогащения антибиотиками.
(A) Схематическое изображение, показывающее образование тройных флуоресцентных нокинов в органоидах рака толстой кишки человека в 2 раундах нацеливания на гены. mScarlet-Blast R был интегрирован в локус CDh2 , кодирующий E-кадгерин, с последующим отбором бластицидином в течение 2 недель для обогащения нокаут-ин органоидами.Последующий ручной отбор дал чистую линию (99,5%, Scarlet+). Во втором раунде нацеливания генов mTurquoise2-Puro R был интегрирован в локус HIST1h3BC в сочетании с интеграциями mNeongreen в локусы LMNA, SEC61B или MAP4. На схемах: положения разрывов Cas9 (D10A) (красные треугольники) и мотивы, прилегающие к протоспейсеру (черная полоса), указаны для каждой нокаутной конструкции, включая используемую гРНК (зеленая стрелка). (B) Графики FACS для органоидов с тройным нокаутом после второго раунда нацеливания и отбора пуромицином в течение 10 дней.Обогащение пуромицином привело к почти полной селекции HIST1h3B-mTQ2-Puro R положительных клеток (фиолетовые и синие популяции объединены). Цифры на графиках FACS указывают частоту (%) нокаутированных популяций во всей целевой клеточной популяции. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ).
(B) Графики FACS для органоидов с тройным нокаутом после второго раунда нацеливания и отбора пуромицином в течение 10 дней.Обогащение пуромицином привело к почти полной селекции HIST1h3B-mTQ2-Puro R положительных клеток (фиолетовые и синие популяции объединены). Цифры на графиках FACS указывают частоту (%) нокаутированных популяций во всей целевой клеточной популяции. Необработанные файлы FCS доступны в FlowRepository (FR-FCM-Z4PJ).
10.1371/journal.pbio.3001527.s007
(EPS)
S8 Рис. Секвенирование по Сэнгеру дифференциальных биаллельных модификаций в опухолевой органоидной линии, полученной от пациента.
Секвенирование по Сэнгеру проводили на поликлональных двуаллельных опухолевых нокаутных линиях, несущих интеграции как mScarlet, так и mNeongreen либо в локусе SEC61B , MAP4 , либо HIST1h3BC . Соединения 5′ и 3′ каждого аллеля с нокаутом показаны для всех 3 условий биаллельного нокаута (см. фиг. 4А). Для HIST1h3BC -mSC сохранялся эндогенный стоп-кодон локуса HIST1h3BC .
10.1371/journal.pbio.3001527.s008
(TIF)
S1 Таблица.Наличие нецелевых эффектов (небольшие вставки и контрольные точки структурных вариаций) в предсказанных in silico нецелевых областях (спейсерные и фланкирующие области) в WGS 3 клональных опосредованных ITPN
SEC61B -mScarlet нокаут-ин органоидных линий.ITPN, парные разрывы внутри транса; WGS, полногеномное секвенирование.
10.1371/journal.pbio.3001527.s009
(XLSX)
Файл S1. Соображения по проектированию и протокол клонирования для одноэтапной сборки вектора-мишени для получения флуоресцентной маркировки генов с использованием никинга или расщепления в транс-паре.
10.1371/journal.pbio.3001527.s015
(DOCX)
Благодарности
Мы благодарим сотрудников лаборатории Snippert за реагенты, предложения и обсуждения. Мы благодарим Маркуса Дж. ван Русмалена за совет по анализу WGS.
Каталожные номера
- 1.
Cai Y, Hossain MJ, Hériché JK, Politi AZ, Walther N, Koch B, et al.
 Экспериментальная и вычислительная основа для динамического белкового атласа деления клеток человека. Природа. 2018;561:411–5.пмид:30202089
Экспериментальная и вычислительная основа для динамического белкового атласа деления клеток человека. Природа. 2018;561:411–5.пмид:30202089 - 2. Клеверс Х. Моделирование развития и болезней с помощью органоидов. Клетка. 2016; 165:1586–97. пмид:27315476
- 3. Шванк Г., Ку Б.К., Сасселли В., Деккерс Дж. Ф., Хео И., Демиркан Т. и др. Функциональное восстановление CFTR с помощью CRISPR/Cas9 в органоидах стволовых клеток кишечника у пациентов с муковисцидозом. Клеточная стволовая клетка. 2013;13:653–658. пмид:24315439
- 4. Андерссон-Рольф А., Мустата Р.С., Меренда А., Ким Дж., Перера С., Грего Т. и др. Одношаговое создание условных и обратимых нокаутов генов.Нат Методы. 2017;14:287–9. пмид:28135257
- 5. Сан Д., Эванс Л., Перроне Ф., Соклева В., Лим К., Резахани С. и др. Набор функциональных генетических инструментов для органоидов, полученных из тканей человека. Элиф. 2021;10:e67886. пмид:34612202
- 6.
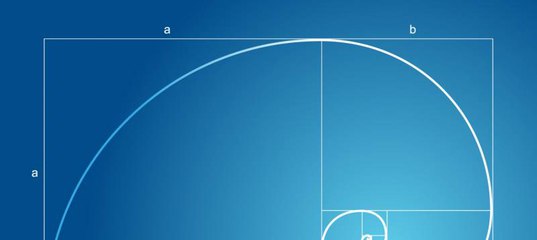 Артегиани Б., Хендрикс Д., Боймер Дж., Кок Р., Чжэн С., Джур И. и др. Быстрое и эффективное создание искусственных человеческих органоидов с использованием независимого от гомологии точного редактирования генома CRISPR-Cas9. Nat Cell Biol. 2020; 22: 321–31. пмид:32123335
Артегиани Б., Хендрикс Д., Боймер Дж., Кок Р., Чжэн С., Джур И. и др. Быстрое и эффективное создание искусственных человеческих органоидов с использованием независимого от гомологии точного редактирования генома CRISPR-Cas9. Nat Cell Biol. 2020; 22: 321–31. пмид:32123335 - 7.Suzuki K, Tsunekawa Y, Hernandez-Benitez R, Wu J, Zhu J, Kim EJ и др. Редактирование генома in vivo с помощью CRISPR/Cas9, опосредованное независимой от гомологии целевой интеграцией. Природа. 2016; 540:144–9. пмид:27851729
- 8. Chen X, Janssen JM, Liu J, Maggio I, T’Jong AEJ, Mikkers HMM и др. В случае транспарных никинг запускает бесшовное редактирование генома без разрезания двухцепочечной ДНК. Нац коммун. 2017;8:657. пмид:28939824
- 9. Рис Х.А., Йе У.Х., Лю Д.Р. Разработка слияний никазы hRad51-Cas9, которые опосредуют HDR без двухцепочечных разрывов.Нац коммун. 2019;10:2212. пмид:31101808
- 10.
Хёдо Т., Рахман М.Л., Карнан С., Ито Т.
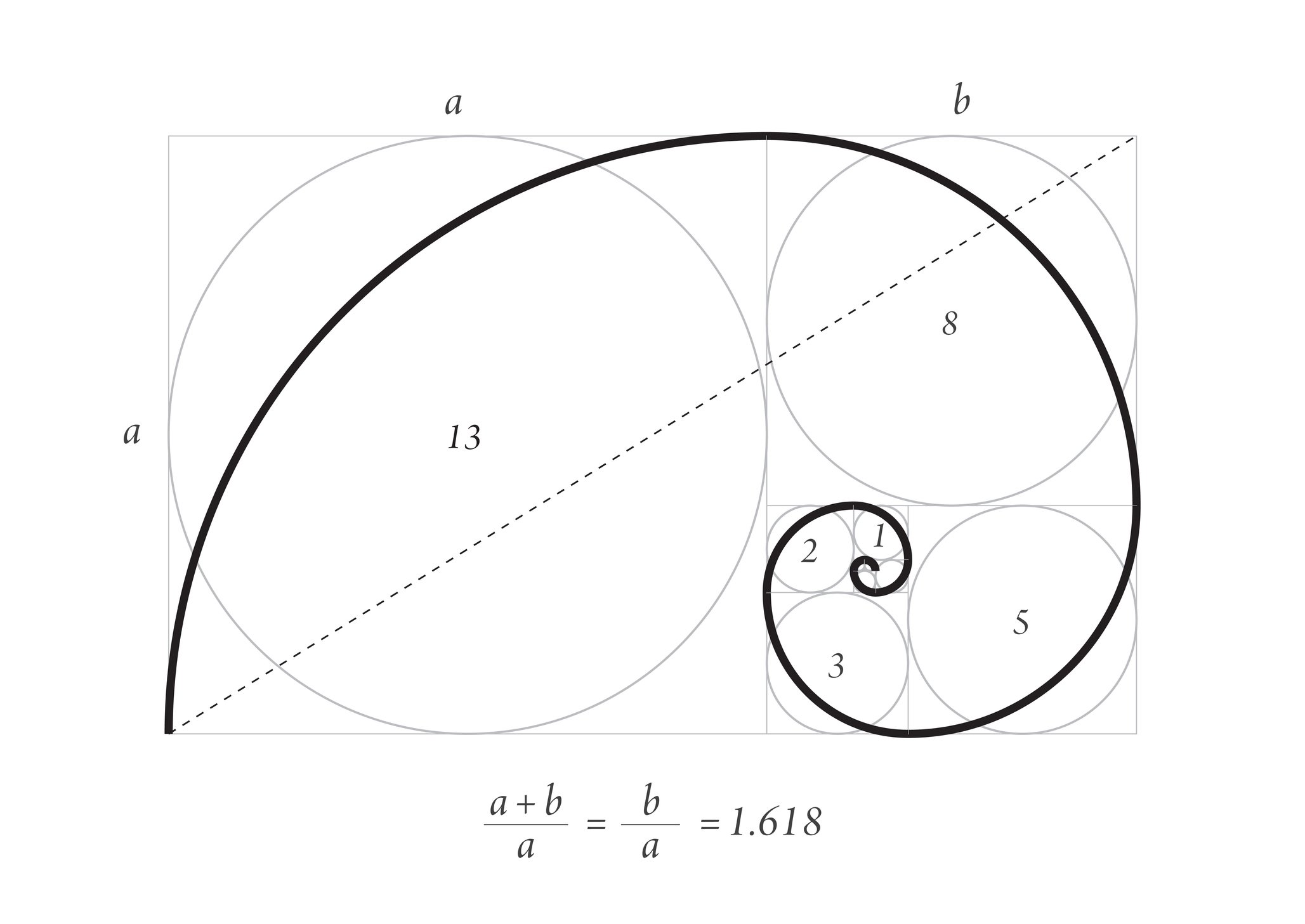 , Тойода А., Ота А. и др. Тандемное парное никирование способствует точному редактированию генома с небольшим вмешательством p53. Cell Rep. 2020;30:1195–1207.e7. пмид:31995758
, Тойода А., Ота А. и др. Тандемное парное никирование способствует точному редактированию генома с небольшим вмешательством p53. Cell Rep. 2020;30:1195–1207.e7. пмид:31995758 - 11. Ран Ф.А., Хсу П.Д., Лин С.И., Гутенберг Дж.С., Конерманн С., Тревино А.Е. и соавт. Двойной разрыв с помощью CRISPR Cas9 под управлением РНК для повышения специфичности редактирования генома. Клетка. 2013; 154:1380–9. пмид:23992846
- 12.Вриенд ЛЕМ, Кравчик ПМ. Инициируемая Ником гомологичная рекомбинация: защита генома, по одной цепи за раз. Восстановление ДНК (Amst). 2017; 50:1–13. пмид:28087249
- 13. Накаде С., Цубота Т., Сакане Ю., Куме С., Сакамото Н., Обара М. и др. Опосредованная микрогомологией интеграция донорской ДНК, зависящая от соединения концов, в клетках и животных с использованием TALEN и CRISPR/Cas9. Нац коммун. 2014;5:5560. пмид:25410609
- 14.
Zhang JP, Li XL, Li GH, Chen W, Arakaki C, Botimer GD и другие.Эффективный точный нокин с донором HDR с двойным разрезом после CRISPR/Cas9-опосредованного расщепления двухцепочечной ДНК.
 Геном биол. 2017;18:35. пмид:28219395
Геном биол. 2017;18:35. пмид:28219395 - 15. Yao X, Wang X, Hu X, Liu Z, Liu J, Zhou H и др. Целевая интеграция на основе соединения концов, опосредованная гомологией, с использованием CRISPR/Cas9. Сотовый рез. 2017; 27:801–14. пмид:28524166
- 16. ван де Ветеринг М., Фрэнсис Х.Е., Фрэнсис Дж.М., Боунова Г., Иорио Ф., Пронк А. и др. Перспективное создание биобанка живых органоидов больных колоректальным раком.Клетка. 2015; 161:933–45. пмид:25957691
- 17. Бринкман Э.К., Чен Т., Амендола М., Ван Стенсел Б. Простая количественная оценка редактирования генома путем разложения следов последовательности. Нуклеиновые Кислоты Res. 2014;42:e168. пмид:25300484
- 18.
Faddah DA, Wang H, Cheng AW, Katz Y, Buganim Y, Jaenisch R. Анализ отдельных клеток показывает, что экспрессия nanog является биаллельной и столь же вариабельной, как и другие факторы плюрипотентности в ESCs мыши. Клеточная стволовая клетка. 2013;13:23–9.
 пмид:23827708
пмид:23827708 - 19. Филипчик А., Гкацис К., Фу Дж., Хоппе П.С., Ликерт Х., Анастассиадис К. и др. Биаллельная экспрессия белка nanog в эмбриональных стволовых клетках мыши. Клеточная стволовая клетка. 2013; 13:12–3. пмид:23827706
- 20. Shaner NC, Lambert GG, Chammas A, Ni Y, Cranfill PJ, Baird MA, et al. Яркий мономерный зеленый флуоресцентный белок, полученный из Branchiostoma lanceolatum. Нат Методы. 2013;10:407–9. пмид:23524392
- 21. Goedhart J, Von Stetten D, Noirclerc-Savoye M, Lelimousin M, Joosen L, Hink MA, et al.Структурно-управляемая эволюция голубых флуоресцентных белков с квантовым выходом 93%. Нац коммун. 2012;3:751. пмид:22434194
- 22. Bindels DS, Haarbosch L, Van Weeren L, Postma M, Wiese KE, Mastop M, et al. mScarlet: яркий мономерный красный флуоресцентный белок для визуализации клеток. Нат Методы. 2017;14:53–6. пмид:27869816
- 23.
Гурская Н.Г., Верхуша В.В., Щеглов А.
 С., Староверов Д.Б., Чепурных Т.В., Фрадков А.Ф. и др. Разработка мономерного фотоактивируемого флуоресцентного белка, переходящего от зеленого к красному, индуцированного синим светом.Нац биотехнолог. 2006; 24:461–5. пмид:16550175
С., Староверов Д.Б., Чепурных Т.В., Фрадков А.Ф. и др. Разработка мономерного фотоактивируемого флуоресцентного белка, переходящего от зеленого к красному, индуцированного синим светом.Нац биотехнолог. 2006; 24:461–5. пмид:16550175 - 24. Ран Ф.А., Хсу П.Д., Райт Дж., Агарвала В., Скотт Д.А., Чжан Ф. Геномная инженерия с использованием системы CRISPR-Cas9. Нат Проток. 2013;8:2281–308. пмид:24157548
- 25. Фуджи М., Матано М., Тошимицу К., Такано А., Миками Ю., Нисикори С. и др. Кишечные органоиды человека сохраняют способность к самообновлению и клеточное разнообразие в условиях культуры, вдохновленной нишами. Клеточная стволовая клетка. 2018;23:787–93. пмид:30526881
- 26.Фуджи М., Матано М., Нанки К., Сато Т. Эффективная генная инженерия кишечных органоидов человека с использованием электропорации. Нат Проток. 2015;10:1474–85. пмид:26334867
- 27.
Ли Х., Хэндсакер Б., Высокер А., Феннелл Т., Руан Дж., Гомер Н. и др. Формат выравнивания/карты последовательностей и SAMtools.
 Биоинформатика. 2009;25:2078–9. пмид:19505943
Биоинформатика. 2009;25:2078–9. пмид:19505943 - 28. Депристо М.А., Бэнкс Э., Поплин Р., Гаримелла К.В., Магуайр Дж.Р., Хартл С. и др. Основа для обнаружения вариаций и генотипирования с использованием данных секвенирования ДНК следующего поколения.Нат Жене. 2011;43:491–501. пмид:21478889
- 29. Блокзейл Ф., Де Лигт Дж., Ягер М., Сасселли В., Реринк С., Сасаки Н. и др. Накопление тканеспецифических мутаций в стволовых клетках взрослого человека в течение жизни. Природа. 2016; 538: 260–4. пмид:27698416
- 30. Джагер М., Блокзейл Ф., Сасселли В., Бойманс С., Янссен Р., Бесселинк Н. и др. Измерение накопления мутаций в отдельных стволовых клетках взрослого человека путем полногеномного секвенирования органоидных культур. Нат Проток. 2018;13:59–78.пмид:29215633
- 31.
Кэмерон Д.Л., Бабер Дж., Шейл С., Папенфусс А.Т., Валле-Инклан Дж.Е., Бесселинк Н. и др. GRIDSS, PURPLE, LINX: расшифровка генома опухоли с помощью комплексного анализа структурных вариаций и числа копий.
 bioRxiv. 2019:781013.
bioRxiv. 2019:781013. - 32. Пэ С, Пак Дж, Ким Дж. Cas-OFFinder: быстрый и универсальный алгоритм, который ищет потенциальные нецелевые участки эндонуклеаз, управляемых РНК Cas9. Биоинформатика. 2014;30:1473–5. пмид:24463181
- 33.Куинлан А.Р., Холл И.М. BEDTools: гибкий набор утилит для сравнения геномных признаков. Биоинформатика. 2010; 26:841–2. пмид:20110278
- 34. Понсиоен Б., Пост Дж. Б., Бьюссан де Амори Дж. Р., Ласкарис Д., ван Иневельд Р. Л., Керстен С. и др. Количественная оценка динамики ERK одиночных клеток в органоидах колоректального рака показывает, что EGFR является усилителем передачи сигналов онкогенного пути MAPK. Nat Cell Biol 2021 234. 2021;23: 377–390. пмид:33795873
Золотое сечение для дизайна логотипов или иконок в Illustrator – tutvid.com
СОЗДАЙТЕ СПИРАЛЬ ЗОЛОТОГО СООТНОШЕНИЯ И ЗНАК-ЛОГОТИП С ПОМОЩЬЮ ЭТОГО РУКОВОДСТВА! | В этом уроке мы создадим прямоугольник золотого сечения с нуля, и вы увидите, насколько простой, увлекательной и полезной может быть эта техника!
В этом уроке Adobe Illustrator мы создадим наш собственный спиральный прямоугольник золотого сечения (последовательность Фибоначчи), а затем воспользуемся изученными методами для создания серии кругов, которые мы можем наложить на эскиз в Illustrator и использовать инструмент Shaper. чтобы быстро отделить и соединить фигуры вместе, чтобы создать простой плоский значок с отрицательным пространством.Эта техника эффективна для очень многих различных дизайнерских приложений, и я думаю, вам это очень понравится! Спасибо за просмотр!
чтобы быстро отделить и соединить фигуры вместе, чтобы создать простой плоский значок с отрицательным пространством.Эта техника эффективна для очень многих различных дизайнерских приложений, и я думаю, вам это очень понравится! Спасибо за просмотр!
Эксклюзивное учебное пособие для сайта Примечания к записи:
Отказ от ответственности: это фактические заметки, которые я использовал для записи этого видео, и они написаны на языке, который вы можете понять или не понять.Надеюсь, вы найдете их полезными или крутыми.
- Создайте прямоугольник 500x500px без заливки и черной обводки
- Создайте эллипс размером 1000×1000 пикселей, такой же, как заливка/обводка прямоугольника
- Включите интеллектуальные направляющие и выровняйте эллипс так, чтобы он пересекал квадрат
- Используйте инструмент Shaper и, удерживая нажатой клавишу Alt/Opt, сотрите выступающие части
- Объект>Разверните и разверните здесь только объект
- Дублируйте сгруппированную фигуру и разделите на 1.
 618 на панели управления и продолжайте дублировать и поворачивать на 90º по часовой стрелке — делайте это, пока у вас не будет 8 групп форм .
618 на панели управления и продолжайте дублировать и поворачивать на 90º по часовой стрелке — делайте это, пока у вас не будет 8 групп форм . - Используйте инструмент «Эллипс», чтобы нарисовать круг, просто продублируйте и разделите круг на 1,618, и вы начнете создавать серию эллипсов, которые будут соответствовать золотому сечению
- Создайте несколько таких изображений и перетащите их с помощью Alt/Opt, чтобы продублировать их и наложить на эскиз, чтобы начать создание графического объекта
- Суетитесь, рисуйте круги и рисуйте голову птицы, создайте тень от щеки и т. д. (не уверен, что это будет объяснено или ускорено для краткости — мы всегда можем сделать еще один урок по этому в будущем.)
Что такое золотое сечение и как его использовать в графическом дизайне
Золотое сечение, также известное как Золотое сечение или Божественная пропорция, представляет собой математическое соотношение 1:1,618 , основанное на последовательности Фибоначчи. Его можно найти в природе (лепестки цветов, семена, скорлупа), в продуктах питания (артишоки, брокколи, ананас) и в анатомии человека.
Его можно найти в природе (лепестки цветов, семена, скорлупа), в продуктах питания (артишоки, брокколи, ананас) и в анатомии человека.
Золотое сечение также можно найти в искусстве (Мона Лиза, Тайная вечеря, Витрувианский человек) и архитектуре (Пирамиды, Акрополь, Нотр-Дам, Тадж-Махал).Применительно к дизайну, фотографии и типографике делает композиции естественными, сбалансированными и эстетически приятными.
КомпанияFolders разработала удобную инфографику, наглядно объясняющую, что такое золотое сечение, с примерами. Мы также включили несколько отличных руководств от дизайнеров Уилла Патерсона и Мохамеда Ахрафа, которые покажут вам, как использовать золотое сечение в графике, логотипе и веб-дизайне. Проверьте это ниже.
Как создать логотип с золотым сечением
Загрузите исходные файлы, используемые в руководстве.
Золотое сечение в композиции и дизайне логотипа
Различные способы использования золотого сечения в вашем дизайне
Золотое сечение Веб-дизайн
Поделитесь этим постом с другом-дизайнером и выскажите свое мнение в комментариях ниже.
Золотое сечение: ключ к идеальному дизайну
Известный во всем мире как Золотое сечение, божественная пропорция, Золотое сечение или Золотое сечение, это математическое чудо, другими словами, значение Фи. На основе знаменитой последовательности Фибоначчи, которая выглядит как 1,1,2,3,5,8,13,21,34… и так далее до бесконечности, где каждое новое число в последовательности является комбинацией двух предыдущих, его соотношение 1,6180 стало известно как идеальная пропорция для создания хорошо сбалансированных и приятных для глаз произведений искусства и архитектуры. На самом деле в любой форме дизайна, от живописи до фотографии и веб-дизайна, золотое сечение стало правильным.
На основе знаменитой последовательности Фибоначчи, которая выглядит как 1,1,2,3,5,8,13,21,34… и так далее до бесконечности, где каждое новое число в последовательности является комбинацией двух предыдущих, его соотношение 1,6180 стало известно как идеальная пропорция для создания хорошо сбалансированных и приятных для глаз произведений искусства и архитектуры. На самом деле в любой форме дизайна, от живописи до фотографии и веб-дизайна, золотое сечение стало правильным.
«Золотое» открытие!
Это началось как средневековая математическая шутка в 12 веке, когда итальянец по имени Леонардо Фибоначчи начал вычислять идеальное разведение кроликов в течение года.Благодаря своим обширным исследованиям он придумал эту серию. Мог ли он знать, что его открытие превратится в волшебную пропорцию, которую можно использовать для создания или проектирования всего, что вы хотите, в идеальном балансе?
Многие исследователи показали нам, что это Отношение 1,6180, вероятно, было основой, используемой египтянами для строительства своих Пирамид, то, как они стоят даже сегодня, невозмутимые и неизменные на протяжении веков, является свидетельством его идеального баланса. Точно так же Парфенон в Афинах, который, как говорят, создан с помощью этого золотого сечения во всех его углах, его колоннах и его вертикальном фасаде, причина, по которой он все еще стоит. В картинах его использовал Леонардо да Винчи в его знаменитой Моне Лизе и Сальвадор Дали в его Таинстве Тайной Вечери; Божественная пропорция, или Фи, или это Золотое сечение, как бы мир ни называл это, можно найти в каждом эстетически совершенном изображении, с которым мы можем столкнуться.
Точно так же Парфенон в Афинах, который, как говорят, создан с помощью этого золотого сечения во всех его углах, его колоннах и его вертикальном фасаде, причина, по которой он все еще стоит. В картинах его использовал Леонардо да Винчи в его знаменитой Моне Лизе и Сальвадор Дали в его Таинстве Тайной Вечери; Божественная пропорция, или Фи, или это Золотое сечение, как бы мир ни называл это, можно найти в каждом эстетически совершенном изображении, с которым мы можем столкнуться.
Божественная пропорция: почему это так?
Его называют божественной пропорцией из-за почти идеальных, безошибочных результатов, которые он дает.Везде, где действует Фи, результат хорошо сбалансирован, гармоничен и так красиво построен, что каждая деталь очевидна со всеми хитросплетениями своего узора, она ярка и в то же время утонченна. Если вы внимательно изучите его, он поразит вас, а если вы просто взглянете на него как наблюдатель, он зачарует. Фотографов учат использовать закон третей для получения изображения с правильными пропорциями, которое не только одинаково во всех углах, но и наиболее приятно для глаз. Один из самых потрясающих снимков всех времен, а также свадебные тренды, которые любят фотографы, основаны на совершенстве Phi.
Один из самых потрясающих снимков всех времен, а также свадебные тренды, которые любят фотографы, основаны на совершенстве Phi.
Поэтому во всем своем совершенстве, точности и «божественности» Золотое сечение является основным правилом всего, что мы создаем.
Даже природа вынуждена этим:
Золотое сечение еще более завораживает, когда оно наблюдается в ходе Природы. Кажется, что все, от Галактик до гор, таких природных явлений, как тайфуны и ураганы, вплоть до деревьев, растений и семенных образований, кажется, что все вещи созданы на основе этого одного соотношения; Фи.Лучшим примером, конечно же, является семя подсолнуха, где они закручиваются в божественной пропорции 1,618, и если мы посчитаем семена любой спирали, число будет последовательностью Фибоначчи.
Ряд Фибоначчи также наблюдается у растений, которые выпускают листья из одного стебля. По мере того, как дерево закручивается вверх по спирали, количество листьев вокруг каждого ствола будет числом Фибоначчи. Действительно поразительно обнаружить, что даже человеческое тело имеет отношение к фи, расстояние пальца от основания до запястья больше, чем расстояние, предшествующее фи.Лепестки цветов имеют такое соотношение, раковины улиток имеют золотую спираль, тела животных демонстрируют аналогичные тенденции, человеческие лица считаются более красивыми, если они ближе к Соотношению в своих пропорциях, даже репродуктивная динамика медоносной пчелы основана на Золотое сечение. Также было отмечено, что схема полета ястреба очень напоминает Золотую Спираль, угол его спуска такой же, как шаг спирали и направление полета. Везде действует Золотое сечение, от спирали Млечного Пути до молекул ДНК, и оно все еще изучается.
Действительно поразительно обнаружить, что даже человеческое тело имеет отношение к фи, расстояние пальца от основания до запястья больше, чем расстояние, предшествующее фи.Лепестки цветов имеют такое соотношение, раковины улиток имеют золотую спираль, тела животных демонстрируют аналогичные тенденции, человеческие лица считаются более красивыми, если они ближе к Соотношению в своих пропорциях, даже репродуктивная динамика медоносной пчелы основана на Золотое сечение. Также было отмечено, что схема полета ястреба очень напоминает Золотую Спираль, угол его спуска такой же, как шаг спирали и направление полета. Везде действует Золотое сечение, от спирали Млечного Пути до молекул ДНК, и оно все еще изучается.
Секрет лучшего эстетического оформления:
Многие люди задавались вопросом, что делает Божественную пропорцию такой идеально сбалансированной, и какое бы изображение, архитектура, графика или графика ни делались с помощью Phi, оно получается очень приятным и красивым. Адриан Бежан, профессор машиностроения Университета Дьюка в Дареме, Северная Каролина, открыл секрет эстетического совершенства золотого сечения.
Адриан Бежан, профессор машиностроения Университета Дьюка в Дареме, Северная Каролина, открыл секрет эстетического совершенства золотого сечения.
Согласно его исследованиям, человеческий глаз ориентирован горизонтально. Так что все, что вычерчено в горизонтальных плоскостях, как Золотой прямоугольник, человеческий глаз не только быстро усваивает, но и ценит его художественность и новаторство, потому что сразу схватывает узоры и хитросплетения изображения. Преднамеренно или случайно, соотношение в Божественной Пропорции лучше всего передается в мозг. Какой бы дизайн, изображение или архитектура ни были основаны на Phi, он получается приятным для глаз результатом, создавая эффект красоты благодаря идеальному балансу и выравниванию.
Проектирование с использованием золотого сечения: простая задача:
В настоящее время золотое сечение также используется для веб-дизайна, и его очень легко включить в любую страницу, которую мы хотим создать. Например, правило третей — это самый быстрый способ создать наилучшее положение для всего контента на вашей странице, поскольку его три горизонтальные и три вертикальные линии поровну делят прямоугольник и автоматически дают вам фокус. Вы можете работать с любой формой, но использование прямоугольника сделает его ближе к золотому прямоугольнику и, таким образом, обеспечит наилучшие результаты.Вы можете настраивать точки, чтобы управлять пикселями на странице, но сохранение значения ближе к 1,6180 даст наилучшие результаты. Таким образом, вы можете легко создать веб-страницу с сертифицированными хорошими результатами.
Например, правило третей — это самый быстрый способ создать наилучшее положение для всего контента на вашей странице, поскольку его три горизонтальные и три вертикальные линии поровну делят прямоугольник и автоматически дают вам фокус. Вы можете работать с любой формой, но использование прямоугольника сделает его ближе к золотому прямоугольнику и, таким образом, обеспечит наилучшие результаты.Вы можете настраивать точки, чтобы управлять пикселями на странице, но сохранение значения ближе к 1,6180 даст наилучшие результаты. Таким образом, вы можете легко создать веб-страницу с сертифицированными хорошими результатами.
Мы также подготовили для вашего рассмотрения короткую, но информативную презентацию, в которой вы найдете ряд примеров дизайна логотипа, которые воплощают универсальную ценность баланса и совершенства. Посмотрите и дайте нам знать о ваших отзывах:
Теперь также очень легко рассчитать все средние значения и медианы с помощью инструментов, которые теперь доступны для макета веб-страницы с использованием золотого сечения. Доступный в виде приложения, он вывел простую задачу на новый уровень. Теперь вы можете легко купить это приложение, и оно проведет вас через все этапы проектирования, а его функции включают в себя «избранное» для повторного использования для повторяющихся задач и режим «щелчка», который позволяет использовать его также в Photoshop. Кроме того, есть несколько фантастических инструментов, которые могут сделать вашу работу намного проще и быстрее, с гарантированным совершенством точности. Возьмем, к примеру, типографский калькулятор . Первоначально вводя некоторую базовую информацию, такую как требуемый размер шрифта или ширина содержимого, его можно легко настроить, чтобы всегда настраивать типографику, чтобы результаты были наилучшими, которые вы хотите.Еще один инструмент, который может помочь в достижении элемента золотого сечения во всех аспектах вашего веб-дизайна, называется PHICALCULATOR . Его простая и точная функция позволяет вычислить значение PHI любого числа за считанные минуты! Это не только экономит ваше время, но и ускоряет процесс и позволяет почувствовать гордость за хорошо выполненную работу.
Доступный в виде приложения, он вывел простую задачу на новый уровень. Теперь вы можете легко купить это приложение, и оно проведет вас через все этапы проектирования, а его функции включают в себя «избранное» для повторного использования для повторяющихся задач и режим «щелчка», который позволяет использовать его также в Photoshop. Кроме того, есть несколько фантастических инструментов, которые могут сделать вашу работу намного проще и быстрее, с гарантированным совершенством точности. Возьмем, к примеру, типографский калькулятор . Первоначально вводя некоторую базовую информацию, такую как требуемый размер шрифта или ширина содержимого, его можно легко настроить, чтобы всегда настраивать типографику, чтобы результаты были наилучшими, которые вы хотите.Еще один инструмент, который может помочь в достижении элемента золотого сечения во всех аспектах вашего веб-дизайна, называется PHICALCULATOR . Его простая и точная функция позволяет вычислить значение PHI любого числа за считанные минуты! Это не только экономит ваше время, но и ускоряет процесс и позволяет почувствовать гордость за хорошо выполненную работу. Для вашего визуального проектирования в качестве программного обеспечения доступен инструмент под названием ATRISE Golden Section, который позволяет вам создавать всю вашу графику и изображения в соответствии с золотым сечением.Вы можете применить изменения во время работы на экране и просмотреть наилучшие возможные результаты.
Для вашего визуального проектирования в качестве программного обеспечения доступен инструмент под названием ATRISE Golden Section, который позволяет вам создавать всю вашу графику и изображения в соответствии с золотым сечением.Вы можете применить изменения во время работы на экране и просмотреть наилучшие возможные результаты.
Вывод:
От сокровенных поворотов природы до спиралевидных галактик, от искусственных зданий до искусства и веб-дизайна — золотое сечение проявляется во всех узорах Вселенной. Ему подчиняются все законы, именно идеальная симметрия в дизайне заставляет все казаться выверенным и гармоничным, соответствующим всему, что его окружает. Его внушающие благоговейный трепет свойства будут продолжать ослеплять умы тех, кто войдет в царство его возможностей, а те, кто стремится разгадать больше его секретов, на каждом шагу будут открывать для себя целый новый мир.Это математическое чудо; предельная, Божественная пропорция.
.
Leave a Comment