Колосок малюнок: Рисунки пшеницы для срисовки (30 фото) • Прикольные картинки и позитив
02.08.1975 


 Разное
Разное
Малюнок колоска пшениці олівцем. Як намалювати пшеницю олівцем поетапно
Ірина Сергіївна Панова
Інтеграція всіх освітніх галузей.
Вік дітей: 5-6 років.
Ціль: Закріплювати знання про жанр живопису пейзаж. Вчити підбирати відповідну кольорову гаму.
Завдання:
Навчальні:
Вчити передавати у малюнку форму, колір та розмір предметів;
Вчити гармонійно розташовувати зображення на аркуші паперу;
Закріпити знання про жанр живопису краєвид.
Розвиваючі:
Розвивати вміння малювати фон, зображуючи небо та землю;
Розвивати бачення естетичної краси у пейзажі.
Виховні:
Виховувати вміння працювати фарбами акуратно.
Попередня робота:Перегляд презентації: «Звідки до нас на стіл хліб прийшов?» Розгляд та малювання по точках колосу, зерна, снопа з колосся.
Обладнання та матеріали:
Репродукції картин із зображенням пшеничних полів, пшеничних колосків

Альбомні листи формату А4, гуаш, пензлі, підставки для пензлів, банки з водою, ганчіркові серветки.
Хід заняття
Вихователь:
Вірші про пшеницю
У пшениці шлях особливий,
Щоб зерна стали здобою
Потрібно сонечком налитися,
Дощової води напитися,
Постояти під небесами
Золоті колоски,
Похитатися на вітрі
І одного ранку
Зберуть у снопи пшеницю,
Відвезуть до села, до станиці
На крилатому млині
зерна перемелються
чиєюсь доброю рукою
Стануть зернятка борошном.
Вихователь: Як ви думаєте, про що ми сьогодні говоритимемо на заняття? (Відповіді дітей)Правильно про хлібі.
Вихователь: Подивіться на картини перед вами, що ви можете про них сказати (Відповіді дітей)
Вихователь
: Правильно це пейзаж Що зображено на краєвиді?Згадаймо, що таке пейзаж? Пейзаж – це жанр мистецтва, що зображує природу чи якусь місцевість (ліс, гай, поле, і т. д.)
д.)
Які жанри мистецтва ви знаєте?
Правильно: Натюрморт і портрет
Вихователь: Давайте розглянемо наш пейзаж, що на ньому намальовано?
(Відповіді дітей).
Діти: Поле. Пшеничне поле. Небо.
Як художник зобразив поле, небо?
Діти: Поле жовтого кольору.
Вихователь: Чому поле жовтого кольору?
Діти: Тому, що воно складається з колосків, А вони жовтого кольору.
Вихователь: Здається, що від картини походить світло Хлопці, давайте ми з вами побудемо художниками та намалюємо краєвид, але щоб почати
Пальчикова гімнастика
Борошно в тісто замісили, стискають і розтискають пальчики.
А з тіста ми зліпили: Приплескують долоньками, «ліплять».
Пиріжки та плюшки, По черзі розгинають пальчики
Здобні ватрушки, починаючи з мізинця.
Булочки та калачі —
Всі ми спекти в печі. Обидві долоні розгортаючи вгору.
Обидві долоні розгортаючи вгору.
Дуже смачно! Гладять живіт.
Вихователь: Спочатку підготуємо фон для малюнка. Як ви думаєте, яке тло ми з вами зображатимемо? (Відповіді дітей).
Вихователь: Так, хлопці, фоном нам послужить блакитне небо пшеничне поле жовтого кольору. Нагадаю вам, що небо має бути прозорим, для цього ми наносимо на папір пензлем чисту воду і додаємо краплю фарби: верхня частина листа – синя фарба, нижня частина листа – жовта. І розганяємо мокрим пензлем фарбу по мокрій поверхні аркуша паперу. (Показ вихователя)Почекаємо трохи, поки лист підсохне.
Фізкультхвилинка.
Булка колосом росла, діти піднімають руки вгору, похитують.
Ними вліво-вправо.
Зерном під жорном була, Вип. Упр. «Годування птахів».
А в печі її спік Плескають однією долонькою по іншій.
Добрий майстер хлібопека! Вип. Упр. «Піднос».
Піднімають великі пальці рук.
Молодці, а зараз починаємо малювати колосок. (Показ вихователя)
Набираємо на пензель фарбу жовтого кольору та малюємо стебло колоса, далі знову набираємо фарбу і шляхом примокування ворсу пензля малюємо зерна в колосі.
Звертаю увагу дітей на те, що основним тлом є поле, а в полезростає безліч колосків. Тому можна
По закінченню малюваннярозглядаємо малюнки.
Рефлексія:
Про який жанр мистецтва ми сьогодні говорили? (Пейзаж)
Що на заняття ми малювали? (пшеничне поле та колоски)
Важко було малювати, що саме? (Відповіді дітей)
Сподобалися вам малюнки? (ТАК)
Подобається вам займатися малюванням? (ТАК)
Вихователь: Ви такі молодці! Дуже гарні пшеничні поля та колосся у вас вийшли, І небо справжнісіньке синє і блакитне!
Оформляємо виставку дитячих робіт.
Публікації на тему:
Конспект інтегрованого заняття з пізнавального розвитку «Хліб — усьому голова» у підготовчій групіЗавдання: — удосконалювати знання про хліб; формувати знання у тому, як виходить борошно; — Активізувати та розширити словниковий запас.
Конспект підсумкового заняття у підготовчій групі у формі гри «Поле чудес»Підсумкове заняття у підготовчій групі у формі гри «Поле чудес» Освітні галузі: «Пізнавальний розвиток», «Мовленнєвий розвиток».
Тема: «Що ми знаємо про Комі краї» Програмний зміст: 1) Систематизувати знання дітей про міста, ріки, тваринний і рослинний світ.
Конспект тематичного заняття у підготовчій групі Тема: «Як хліб на стіл прийшов»Ціль: формування уявлення дітей про працю дорослих; показувати результати праці та її суспільну значимість, вчити дбайливо ставитися.
Конспект заняття з декоративного малювання у підготовчій групі.Тема: «Півник із сім’єю» (набір сувенірів за мотивами Гжелі). Завдання: 1. Прикрашати елементи розпису гжельського предмети складного контуру.
Завдання: 1. Прикрашати елементи розпису гжельського предмети складного контуру.
Конспект заняття з навколишнього світу у підготовчій групі «Хліб — усьому голова»Конспект заняття з навколишнього світу у підготовчій групі
Урок присвячується всім ботанам та ненажерам. Ви побачите, як малювати пшеницю олівцем поетапно. Звичайно малювати ціле поле я не збираюся, зате покажу кілька колосків:
Пшениця це їстівна трава. З неї роблять багато смакот: хліб, макарошки, солодощі, пиво та інші шляхетні напої. Тож її люблять і дорослі, і діти. Щоправда, вирощувати, молоти, і обробляти ніхто не любить, але це вже зовсім інша розмова. Щоб зобразити колосок пщениці треба багато терпіння та уважності. Особливої техніки зображення тут немає, тим більше не потрібно дотримуватись пропорцій. Видів і сортів пшениці настільки багато, що вона сама навіть не знає, якої виросте. З іншого боку, треба зробити красиво, повторюйте за мною.
Як намалювати пшеницю олівцем поетапноКрок перший. Робимо малюнок, як очерету.
Робимо малюнок, як очерету.
Крок другий. Додаємо кругленькі форми, схожі на вербу.
Крок третій. Поправляємо контури, додаємо вусики.
Олена Баранова
«Колосок». Конспект НОД з ГО «Художньо-естетичний розвиток» (малювання) у старшій групі компенсуючого виду
Проектувальний компонент.
Тема: « Колосок» .
Інтеграція освітніх галузей: художньо-естетичний розвитокфізичне розвиток, мовленнєве розвиток, пізнавальне розвиток, соціально-комунікативне розвиток.
Ціль: навчання дітей малювання колосканетрадиційною технікою «примокування».
Завдання:
Корекційно-освітні: розширити знання дітей про колоску, вчити малювати колосок гуашевою фарбою, передаючи особливості його будови, закріпити навички малюванняза допомогою примокування ворсу пензлика, активувати в мовленні дітей прикметник (золотистий, іменник (остя).
Корекційно-виховні: виховувати художній смак, виховувати бажання допомагати іншим,
Корекційно-розвиваючі: розвивати слухову увагу, мислення, дрібну моторику, розвивати почуття ритму, розвиватиестетичне сприйняття.
Обладнання та матеріали:
Театр м’якої іграшки (півник, мишенята, сонечко, іграшкова хатинка, іграшковий віник, натуральний пшеничний колосок, мольберт, альбомні листи формату А4, гуаш золотиста, пензлі двох видів (№6 і №2, стаканчики з водою, серветки (на кожну дитину, коровай).
Дитяча діяльність:
ігрова – гра «Каравай», фізхвилинка з мовним супроводом; комунікативна — питання-відповіді, гра «Каравай»; образотворча- малювання; музична — спів пісні «Сонечко», «Каравай», музично-ритмічні рухи у фізхвилинці «В землю сонечко потрапило»; пізнавально-дослідницька-розгляд колоска, рухова-оволодіння основними рухами у грі «Каравай», сприйняття художньоїлітератури та фольклору: слухання казки
Попередня робота: читання української казки « Колосок» , розгляд колосків, розгляд ілюстрацій, перегляд мультфільму « Колосок» , розучування пісні «Сонечко», розучування фізкультурної хвилинки «В землю сонечко потрапило», гра в «Каравай».
Організаційний компонент:
Підготовка до заняття:
5 баночок із золотистою гуашшю, 5 альбомних аркушів паперу, 5 серветок, 10 пензликів, 5 стаканчиків із водою.
Проведення занять відповідно до СанПіН 2.4.1.3049-13 та здоров’язберігаючими технологіями.
Тривалість безперервної безпосередньо освітньої діяльності — не більше 25 хвилин, у середині часу, відведеного на НОД, проведення фізкультхвилинки.
Метод:
1) Наочний,
2) Словесний,
3) Практичний,
4) Ігровий.
Прийоми: загадка, гра, показ, пояснення
Мотивація:
Театр м’якої іграшки (півник, мишенята).
Хід заняття:
Діти, подивіться до нас прийшли гості. Давайте з ними привітаємось.
Діти вітаються.
Можна нашим гостям бути у нас.
Діти дозволяють.
Сьогодні хочу розповісти Вам казку. Бажаєте її послухати?
Діти погоджуються.
Добре. Тоді займайте будь-які вільні місця.
Жили-були два мишеня, Круть і Верть, та півник Голосисте Шийка. Мишенята тільки й знають, що танцюють та грають, а півник зранку встане, всіх піснею розбудить.
Давайте, хлопці, допоможемо нашому півнику розбудити сонечко.
Діти співають пісеньку та виконують рухи :
Сонечко, сонечко, (зображення руками сонечка)
Виглянь у віконце. (зображення руками віконця)
На хлопців подивися, (руки в сторони)
Ти дітям посвіти! (зображення руками ліхтариків)
Виглядає сонечко.
Півник:
Дякую, хлопці, за допомогу! Піду я на подвір’я підмітати.
Ой, що знайшов! Але спочатку, відгадайте загадку:
Золотист він і вусати,
У ста кишенях сто хлопців.
Здогадалися?
Відповіді дітей.
Правильно, це колосок. Колір золотистий. Ось вуса, ось кишеньки, а ось і хлопці зернятка (Показує). А ще, що є у колоска?
Відповіді дітей.
Покличу я мишенят.
Крути, Верть! Дивіться, що знайшов!
Це колосок! Потрібно його обмолотити!
А хто його обмолотить?
Тільки не я!
Тільки не я!
Гаразд, я обмолочу!
Хлопці, давайте пограємося з Вами, згадаємо, що потрібно, щоб виросли колоски, а потім я обмолочу колосок.
Фізкультурна хвилина «У землю зернятко потрапило» (під музику музичного керівника):
1. У землю зернятко потрапило.
(Діти сідають навпочіпки, притискають голову до колін і закривають її руками)
2. Проростати на сонці стало.
(Діти піднімають руки вгору над головою, примовляючи:
«Золоте сонечко, ти зігрій нам зернятко!»
«Дощику, дощику, поливай! Буде хліба врожай!)
(Діти витягують руки вперед, долонями вгору та «бризкають»на себе водою.)
3. Дощ землю поливав, і паросток підростав!
(Діти повільно встають).
4. До світла та тепла тягнувся (руки в сторони)
5. І красенем обернувся (руки вперед).
І красенем обернувся (руки вперед).
6. Золоті колоски на вітрі гойдаються,
(Діти встають на шкарпетки, піднімають прямі руки вгору, кисті рук опущені вниз).
7. Низько нахиляються.
(Діти махають піднятими вгору руками та примовляють:
«Встиг урожай! Приїдь, прибирай!
(Нахили вниз)
Спасибі хлопці! А де мій колосок? У мене сьогодні іменини і я хотів спекти коровай!
Вихователь:
Не засмучуйся, півнику. Хлопці, давайте допоможемо півня, намалюємо йому колоски.
Діти погоджуються та сідають за столи.
Давайте пригадаємо, як треба сидіти, коли малюємо: «Сяду прямо не зігнуся, за роботу я візьмуся!»
Вихователь малює на мольберті та коментує.
Беремо велику пензлик в руку, занурюємо хвостик пензлика у воду, зайву крапельку віджимаємо об край стаканчика, набираємо гуаш на хвостик, занурюємо тільки хвостик, щоб пензлик не забруднилася. Малюємо стебло знизу вгору, знаходимо середину стебла і ставимо крапку, від цієї точки вгору по різні боки стеблинки малюємо кишеньки, в яких знаходяться зернятка технікою «примокування». Згадуємо, як ми це робимо. Прикладаємо хвостик пензлика до паперу і відразу ж забираємо. Шлеп-шлеп-шлеп. Виходить рівний, гарний відбиток. І останній раз ляпнемо пензликом на самому верху колоска. Тепер беремо тонкий пензлик і малюємо вусики у колоска, акуратно між кишеньками. Вони називаються кості. А хто вже намалював гостя, можна, можливо намалювати листочки.
Згадуємо, як ми це робимо. Прикладаємо хвостик пензлика до паперу і відразу ж забираємо. Шлеп-шлеп-шлеп. Виходить рівний, гарний відбиток. І останній раз ляпнемо пензликом на самому верху колоска. Тепер беремо тонкий пензлик і малюємо вусики у колоска, акуратно між кишеньками. Вони називаються кості. А хто вже намалював гостя, можна, можливо намалювати листочки.
Діти малюють (самостійна діяльність дітей, вихователь допомагає при необхідності індивідуально кожній дитині та хвалить їх).
Діти, покладіть свої малюнки на один стіл! Нехай вони висохнуть!
Діти, що ми з Вами сьогодні робили?
Відповіді дітей.
Що є у колоска?
Відповіді дітей.
Як називаються вусики у колоска?
Відповіді дітей.
Півник милується роботами дітей, хвалить хлопців та дякує їм за допомогу!
Сюрпризний момент:
Вдаються мишенята і приносять коровай!
Півник, вітаємо тебе з іменинами! Це ми взяли твій колосок, обмолотили його, віднесли на млин, намололи муки, витопили піч, замісили тісто і спекли коровай для тебе!
Дякую, мишенята!
Хлопці, давайте привітаємо півня і заспіваємо йому коровай!
Діти вітають півня та співають коровай (під музику музичного керівника):
Як на Петіни іменини спекли ми коровай:
(Діти водять хоровод навколо півника)
Ось такої висоти,
(Піднімають зімкнуті руки вгору)
Ось такої нижини,
(діти сідаючи навпочіпки, опускають руки)
Ось такої ширини.
(Діти розходяться на ширину витягнутих рук, розтягуючи коло)
Коровай, коровай, кого любиш вибирай!
(півник обирає дітей, обходячи їх по колу)
Я, правду кажу, люблю всіх, а от (ім’я обраної дитини)краще за всіх!
(Показує на одного з дітей у колі).
Вибрана дитина та півник танцюють у колі. Гра повторюється, доки всі діти не станцюють.
Півник і мишенята короваєм пригощають дітей і прощаються з ними.
Рефлексивний компонент:
Мета та завдання було досягнуто, діти були активні, їм все сподобалося. Продуктивний вид діяльності – малюванняможна було замінити на ліплення.
Література:
1. Ликова І. А Образотворча діяльність у дитсадку.
2. Українська казка « Колосок» .
3. Малова В. В. конспектизанять із духовно- моральному вихованню дошкільнят на матеріалі російської народної культури.
4. Аверіна І. Є. Фізкультурні хвилини у дитячому садку.
Тематична добірка ігор та вправ для дітей раннього віку на тему «Колосок»
Цілі:
Продовжуватиме вчити дітей уважно слухати казку. Дивитись показ настільного театру, емоційно сприймати зміст.
Ознайомити дітей із казкою «Колосок.
Збагачувати словник дітей на тему.
Формувати стійкі уявлення про величину, кількість, колір, геометричні фігури.
Вправляти в ліпленні, наклеюванні, малюванні пальчиками, пензликами та олівцями.
Розвивати вміння повторювати рухи за вихователем.
Розвивати мислення, моторику, зорове та слухове зосередження, координацію рухів.
Виховувати інтерес до казок.
Обладнання:
Іграшки-герої казки «Колосок», пшеничні колоски, зерна пшениці, борошно, тісто.
Кольорові силуети півня, будиночок, мишенят, колоска, малюнок тло.
Картинка-заготівля «колосок» для малювання пальчиків, пальчикові фарби, олівці жовтого кольору.
Пластилін жовтий.
Картинка-заготівля «Колоски без стебел», жовта гуаш, пензлі.
Зображення фону з зображенням лісу, мишеня, лисиці, два гриби під деревом; вирізані з картону листочки різної величини.
Зображення фону з зображенням колоска з овалів, овали, вирізані з жовтого картону, лічильні палички.
Колоски, вирізані із щільного картону, різнокольорові прищіпки.
Ґудзики різного кольору та величини, картинка із зображенням пирога з кольоровими колами, кольором та величиною, що відповідають ґудзикам.
Картинка-заготівля із зображенням підносів різної форми, силуетні картинки круглої булочки, овальної плетінки, квадратного пирога. Клей, мак, кунжут, пластилін червоного кольору.
Вирізані з кольорового картону серветки, блюдця, чашки, ложки.
Булочки. Сіто. Горох (не лущений).
Аудіозапис: Топ-топ.
Хід заняття:
Гра-вітання «Наші розумні головки»
Наші розумні головки
Думатимуть багато, спритно.
Вуха слухатимуть,
Ротик чітко говорити.
Ручки плескатимуть,
Ніжки тупатимуть.
Спинки випрямляються,
Один одному посміхаємось.![]()
Сьогодні на нас чекає зустріч із казкою.
Показ настільного театру «Колосок»
Жили-були два мишеня, Круть і Верть, та півник Голосна шийка. Мишенята тільки й уміли співати та танцювати, крутитися і крутитися. Півник же рано-вранці піднімався, спершу всіх піснею будив, а потім за роботу приймався.
Якось підмітав півник двір і побачив на землі пшеничний колосок. Покликав півник мишенят:
— Крути та Верть, подивіться, що я знайшов!
Прибігли мишенята і кажуть:
— Треба його обмолотити.
— А хто молотитиме? — Запитав півник.
— Тільки не я! — пропищав Круть.
— Тільки не я! – пропищав Верть.
— Гаразд, — сказав півник, — я обмолочу.
І взявся до роботи. Мишенята тим часом затіяли гру в лапту.
Закінчив півник молотити і крикнув:
— Гей, Круть, гей, Верть, подивіться скільки я зерна намолотив!
Прибігли мишенята і запищали в один голос:
— Тепер треба зерно на млин нести, муки намолоти.
— А хто понесе? — Запитав півник.
— Тільки не я! — відповів Круть.
— Тільки не я! – відповів Верть.
— Гаразд, — сказав півник, — я знесу зерно на млин.
Взяв собі на плечі мішок і пішов. А мишенята затіяли гру в чехарду. Стрибають один через одного, веселяться.
Повернувся півник із млина і покликав мишенят:
— Сюди, Круть, сюди, Верть! Я муку приніс.
Прибігли мишенята, дивляться, не натішаться:
— Ай та півник! Ай та молодець! Тепер треба тісто замісити та хліб пекти.
— Хто міситиме? — Запитав півник.
А мишенята знову своє:
— Тільки не я! — запищав Круть.
— Тільки не я! – запищав Верть.
Подумав, подумав півник і каже:
— Видно мені доведеться.
Замісив півник тісто, натяг дров, затопив піч. Як піч розтопилася, посадив у неї хліб.
Мишенята теж часи не втрачають: танцюють, та пісні співають.
Спекся хліба, півник вийняв, поставив на стіл, а мишенята тут як тут. І звати їх не довелося.
— Ох і зголоднів я! — пропищав Круть.
— Ох, як їсти хочеться! – пропищав Верть.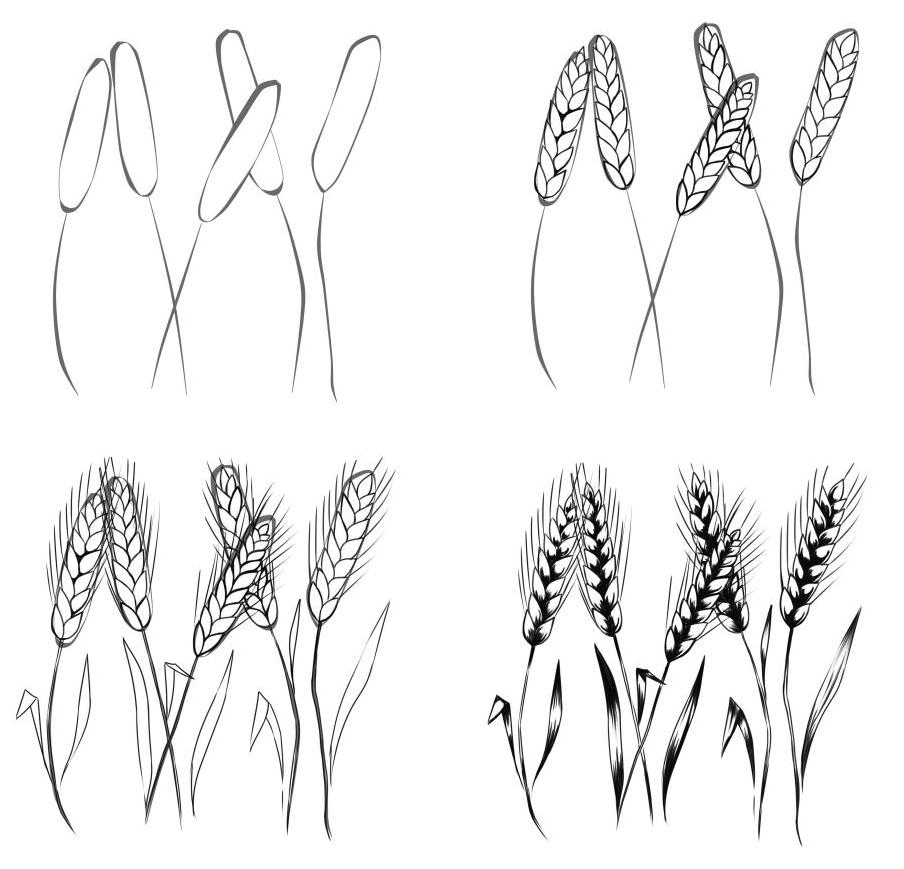
Швидше сіли за стіл. А півник їм каже:
— Зачекайте, зачекайте! Ви мені спершу скажіть, хто знайшов колосок?
— Ти знайшов! — голосно закричали мишенята.
— А хто колосок обмолотив? — знову спитав півник.
— Ти обмолотив! — тихіше сказали обидва.
— А хто зерно на млин носив?
— Теж ти, — зовсім тихо відповіли Круть та Верть.
— А тісто хто місив? Дрова носив? Пекти топив, хліб хто пек?
— Усі ти. Все ти, — трохи чутно пропищали мишенята.
– А ви що робили?
Не знаю, що сказати мишенята. Стали Круть та Верть вилазити з-за столу, а півник їх не тримає.
Нема за що таких ледарів і ледарів хлібом пригощати!
Дидактична гра «Виклади казку на зображенні»
Знайдіть на картинці місце для хатинки, для півня та мишенят. Покладіть колосок під деревом.
Дидактична гра «Сховай мишеня від лисиці»
Вирішили мишенята виправитися — самим зробити пиріг та півня почастувати. Одне мишеня тісто замісило, а інше в ліс пішло — грибів для пирога набрати.
Прийшло мишеня в ліс, побачило гриби. А ви бачите на картинці гриби? Де вони? Під деревом. А скільки грибів під деревом? Два гриби.
Але не пощастило мишеняті, тільки хотів гриби зірвати – попався на очі лисиці. Швидше підберіть лист, відповідного розміру, щоб сховати мишеня від лисиці.
Дидактична гра «Склади колосок»
З яких геометричних фігур складатимемо колосок? З овалів. Скільки колосків у вас на зображенні? Один колосок. А скільки зернят-овалів знадобиться для одного колоска? Багато. Викладіть овали на картинку. А тепер лічильними паличками зробіть вусики колоску.
Малювання «Колосок»
Намалюйте олівцем жовтого кольору вусики колоску. Залишіть відбитки подушечки пальчика на місці зерен у колоску.
Ліплення «Колосок»
З пластиліну жовтого кольору прямим розкочуванням скачайте ковбаску і прикладіть до картону, злегка придавіть її зверху долонею. Покладіть паличку-зубочистку зверху на ковбаску-колосок (вздовж) і вдавіть. А тепер беріть зерна пшениці та вдавлюйте з двох боків від палички.
А тепер беріть зерна пшениці та вдавлюйте з двох боків від палички.
Дидактична гра «Чого не стало»
Перед дітьми викладаються колосок, булочка, тісто. Накриваються хусткою. Один предмет ховається. Потім хустку знімають і дітям пропонується сказати, якого предмета не стало.
Музично-динамічна пауза «Топ-топ»
Діти повторюють рухи під музику за показом вихователя.
Гра з прищіпками «Колосок»
Зробіть із прищіпок колоскам вусики.
Гра з гудзиками «Пиріг»
Розкладіть гудзики на круги відповідного кольору та розміру.
Образотворча діяльність «Пироги та булочки»
Наклейте на підноси пиріг та булки. Підбирайте піднос, що підходить за формою.
Посиплемо круглу булочку маком (попередньо змастивши клеєм), а овальну булочку — кунжутом. А ось пиріг зробимо з вишнями – наліпіть із пластиліну червоного кольору кола-вишні на пиріг.
Дидактична гра «Чайний сервіз»
Підберіть до кожного блюдця чашку і ложку, покладіть на серветки відповідного кольору.
Малювання фарбами «Стеблі колосків»
Намалюйте пензликом стебла колосків.
Вправа «Просіюємо муку через сито»
Дітям пропонується ложкою покласти борошно в сито і просіяти борошно над тацею.
Ліплення із солоного тіста «Пиріг»
Діти роблять із солоного тіста кулю, розплющують її в коржик, розкочують тонкі ковбаски і викладають сіточкою на пиріг. У вільні місця вдавлюють горошини.
Дидактична вправа «Що спочатку, що потім»
Спочатку півник знайшов колосок, покажіть колоски.
Потім із колоска дістав зерна. Покажіть зерна.
З зерен зробив муку. Покажіть муку.
З борошна зробив тісто. Покажіть тісто.
Із тіста зробив пиріг. А ось пиріг покажу вам я. І пропоную спробувати його.
Проводиться дегустація пирога.
Урок присвячується всім ботанам та ненажерам. Ви побачите, як малювати пшеницю олівцем поетапно. Природно малювати ціле я не збираюся, зате покажу кілька колосків:
Пшениця це їстівна трава. З неї роблять багато смакот: хліб, макарошки, солодощі, пиво та інші шляхетні напої. Тож її люблять і дорослі, і діти. Щоправда, вирощувати, молоти, і обробляти ніхто не любить, але це вже зовсім інша розмова. Щоб зобразити колосок пщениці треба багато терпіння та уважності. Особливої техніки зображення тут немає, тим більше не потрібно дотримуватись пропорцій. Видів і сортів пшениці настільки багато, що вона сама навіть не знає, якої виросте. З іншого боку, треба зробити красиво, повторюйте за мною.
З неї роблять багато смакот: хліб, макарошки, солодощі, пиво та інші шляхетні напої. Тож її люблять і дорослі, і діти. Щоправда, вирощувати, молоти, і обробляти ніхто не любить, але це вже зовсім інша розмова. Щоб зобразити колосок пщениці треба багато терпіння та уважності. Особливої техніки зображення тут немає, тим більше не потрібно дотримуватись пропорцій. Видів і сортів пшениці настільки багато, що вона сама навіть не знає, якої виросте. З іншого боку, треба зробити красиво, повторюйте за мною.
Крок перший. Робимо малюнок, як очерету. Крок другий. Додаємо кругленькі форми, схожі на . Крок третій. Поправляємо контури, додаємо вусики. Крок четвертий. Додамо штрихування та великі вусики. Готово: Спробуйте зобразити такі рослини.
Рекомендуємо також
Тематическая подборка игр на тему «Колосок»
Тематическая подборка игр и упражнений для детей раннего возраста на тему «Колосок»
Автор занятия: Давыдова Светлана Алексеевна.
Цели:
Продолжать учить детей внимательно слушать сказку. Смотреть показ настольного театра, эмоционально воспринимать содержание.
Познакомить детей со сказкой «Колосок.
Обогащать словарь детей по теме.
Формировать устойчивые представления о величине, количестве, цвете, геометрических фигурах.
Упражнять в лепке, наклеивании, рисовании пальчиками, кисточками и карандашами.
Развивать умение повторять движения за воспитателем.
Развивать мышление, моторику, зрительное и слуховое сосредоточение, координацию движений.
Воспитывать интерес к сказкам.
Оборудование:
Игрушки-герои сказки «Колосок», пшеничные колоски, зерна пшеницы, мука, тесто.
Цветные силуэты петушка, домика, мышат, колоска, картинка-фон.
Картинка-заготовка «колосок» для пальчикового рисования, пальчиковые краски, карандаши желтого цвета.
Пластилин желтого цвета.
Картинка-заготовка «Колоски без стеблей», желтая гуашь, кисти.
Картинка-фон с изображением леса, мышонка, лисы, двух грибов под деревом; вырезанные из картона листики разной величины.
Картинка-фон с изображением колоска из овалов, овалы, вырезанные из желтого картона, счетные палочки.
Колоски, вырезанные из плотного картона, разноцветные прищепки.
Пуговицы разного цвета и величины, картинка с изображением пирога с цветными кругами, цветом и величиной соответствующии пуговицам.
Картинка-заготовка с изображением подносов разной формы, силуэтные картинки круглой булочки, овальной плетенки, квадратного пирога. Клей, мак, кунжут, пластилин красного цвета.
Вырезанные из цветного картона салфетки, блюдца, чашки, ложки.
Булочки. Сито. Горох (не лущеный).
Аудиозапись: «Топ-топ».
Ход занятия:
Игра-приветствие «Наши умные головки»
Наши умные головки
Будут думать много, ловко.
Ушки будут слушать,
Ротик четко говорить.
Ручки будут хлопать,
Ножки будут топать.
Спинки выпрямляются,
Друг другу улыбаемся.
— Сегодня нас ожидает встреча со сказкой.
Показ настольного театра «Колосок»
— Жили-были два мышонка, Круть и Верть, да петушок Голосистое горлышко. Мышата только и умели петь да плясать, крутиться и вертеться. Петушок же рано утром поднимался, сперва всех песней будил, а потом за работу принимался.
Однажды подметал петушок двор и увидел на земле пшеничный колосок. Позвал петушок мышат:
— Круть и Верть, посмотрите, что я нашел!
Прибежали мышата и говорят:
— Нужно его обмолотить.
— А кто будет молотить? — спросил петушок.
— Только не я! — пропищал Круть.
— Только не я! — пропищал Верть.
— Ладно, — сказал петушок, — я обмолочу.
И принялся за работу. Мышата тем временем затеяли игру в лапту.
Закончил петушок молотить и крикнул:
— Эй, Круть, эй, Верть, посмотрите сколько я зерна намолотил!
Прибежали мышата и запищали в один голос:
— Теперь надобно зерно на мельницу нести, муки намолоть.
— А кто понесет? — спросил петушок.
— Только не я! — ответил Круть.
— Только не я! — ответил Верть.
— Ладно, — сказа петушок, — я снесу зерно на мельницу.
Взвалил себе на плечи мешок и пошел. А мышата затеяли игру в чехарду. Прыгают друг через друга, веселятся.
Вернулся петушок с мельницы и позвал мышат:
— Сюда, Круть, сюда, Верть! Я муку принес.
Прибежали мышата, смотрят, не нарадуются:
— Ай да петушок! Ай да молодец! Теперь нужно тесто замесить да хлеб печь.
— Кто будет месить? — спросил петушок.
А мышата опять свое:
— Только не я! — запищал Круть.
— Только не я! — запищал Верть.
Подумал, подумал петушок и говорит:
— Видно мне придется.
Замесил петушок тесто, натаскал дров, затопил печь. Как печь истопилась, посадил в нее хлеб.
Мышата тоже времни не теряют: пляшут, да песни поют.
Испекся хлебушек, петушок вынул, поставил на стол, а мышата тут как тут. И звать их не пришлось.
И звать их не пришлось.
— Ох и проголодался я! — пропищал Круть.
— Ох, как есть хочется! — пропищал Верть.
Скорее сели за стол. А петушок им говорит:
— Подождите, подождите! Вы мне сперва скажите, кто колосок нашел?
— Ты нашел! — громко закричали мышата.
— А кто колосок обмолотил? — снова спросил петушок.
— Ты обмолотил! — потише сказали оба.
— А кто зерно на мельницу носил?
— Тоже ты, — совсем тихо ответили Круть и Верть.
— А тесто кто месил? Дрова носил? Печь топил, хлеб кто пек?
— Все ты. Все ты, — чуть слышно пропищали мышата.
— А вы что делали?
Не знаю, что сказать мышата. Стали Круть и Верть вылезать из-за стола, а петушок их не удерживает.
Не за что таких лодырей и лентяев хлебом угощать!
Дидактическая игра «Выложи сказку на картинке»
— Найдите на картинке место для избушки, для петушка и мышат. Положите колосок под деревом.
Дидактическая игра «Спрячь мышонка от лисы»
— Решили мышата исправиться — самим сделать пирог и петушка угостить. Один мышонок тесто замесил, а другой в лес пошел — грибов для пирога набрать.
Пришел мышонок в лес, увидал грибы. А вы видите на картинке грибы? Где они? Под деревом. А сколько грибов под деревом? Два гриба.
Но не повезло мышонку, только хотел грибы сорвать — попался на глаза лисе. Скорее подберите листик, подходящего размера, чтобы спрятать мышонка от лисы.
Дидактическая игра «Сложи колосок»
— Из каких геометрических фигур будем складывать колосок? Из овалов. Сколько колосков у вас на картинке? Один колосок. А сколько зернышек-овалов понадобится для одного колоска? Много. Выложите овалы на картинку. А теперь счетными палочками сделайте усики колоску.
Рисование «Колосок»
— Нарисуйте карандашом желтого цвета усики колоску. Оставьте отпечатки подушечки пальчика на месте зерен в колоске.
Лепка «Колосок»
— Из пластилина желтого цвета прямым раскатыванием скатайте колбаску и приложите ее к картону, слегка придавите ее сверху ладонью. Положите палочку-зубочистку сверху на колбаску-колосок (вдоль) и вдавите. А теперь берите зерна пшеницы и вдавливайте с двух сторон от палочки.
Дидактическая игра «Чего не стало»
Перед детьми выкладываются колосок, булочка, тесто. Накрываются платком. Один предмет прячется. Затем платок снимается и детям предлагается сказать, какого предмета не стало.
Музыкально-динамическая пауза «Топ-топ»
Дети повторяют движения под музыку по показу воспитателя.
Игра с прищепками «Колосок»
— Сделайте из прищепок колоскам усики.
Игра с пуговицами «Пирог»
— Разложите пуговицы на круги подходящего цвета и размера.
Изобразительная деятельность «Пироги и булочки»
— Наклейте на подносы пирог и булки. Подбирайте поднос, подходящий по форме.
Посыплем круглую булочку маком (предварительно смазав клеем), а овальную булочку — кунжутом. А вот пирог сделаем с вишнями — налепите из пластилина красного цвета круги-вишни на пирог.
Дидактическая игра «Чайный сервиз»
— Подберите к каждому блюдцу чашку и ложку, положите их на салфетки подходящего цвета.
Рисование красками «Стебли колосков»
— Нарисуйте кисточкой стебли колоскам.
Упражнение «Просеиваем муку через сито»
Детям предлагается ложкой положить муку в сито и просеять муку над подносом.
Лепка из соленого теста «Пирог»
Дети делают из соленого теста шар, расплющивают его в лепешку, раскатывают тонкие колбаски и выкладывают сеточкой на пирог. В свободные места вдавливают горошины.
Дидактическое упражнение «Что сначала, что потом»
— Сначала петушок нашел колосок, покажите колоски.
Затем из колоска достал зерна. Покажите зерна.
Из зерен сделал муку. Покажите муку.
Покажите муку.
Из муки сделал тесто. Покажите тесто.
Из теста сделал пирог. А вот пирог покажу вам я. И предлагаю попробовать его.
Проводится дегустация пирога.
Скачать конспект игрового занятия для детей 2-3 года жизни, тема: «Сказка «Колосок»
схема в’язання візерунка на спицях по колу
У статті ви зможете знайти опис, фотографії, про те, як зв’язати спицями візерунок — азіатський колосок. Так само ви побачите докладний майстер-клас на цю тему у відео.
Зараз для в’язання тієї чи іншої речі можна знайти безліч дуже красивих і оригінальних візерунків. Азіатський колосок використовують багато умільці. І не дарма, адже такий об’ємний візерунок чудово прикрасить будь теплий одяг, особливо він добре виглядає на кардіганах. Правда, пряжі на ньому заощадити не вийде. Так і вільним часом треба запастись, бо в’язання тривалий процес. Далі детальніше вивчимо, як пов’язати на спицях азіатський колосок, в основі якого лежить нехитрий ажурний візерунок.![]()
Азіатський колосок спицями — ледачий візерунок для початківців
Візерунок являє собою досить-таки об’ємний колосок. Нижче на зображенні представлений приклад азіатського колоска. Його використовують для в’язання, як вже згадувалося вище, кардиганів, судин, пледів, пальто, светри, головних уборів. Отримані вироби зігрівають в холоди і відмінно виглядають на модницах. Зв’язати гарний об’ємний малюнок можна різними варіантами.
Азіатський колосок на спицяхДаний варіант підходить тим, хто не дуже добре розбирається в техніці створення візерунка. Об’ємний ледачий малюнок можна пов’язати двома методами. Перший варіант: слід в’язати петлі частинами ряд за рядом, після їх закривають, потім набирають знову.
Якщо в’язати другим способом, то в першому ряду виконується відразу набір, а потім закриття петельок. Візерунок ж в’яжеться окремо.
За другим методом, в 1-му рядку необхідно набрати і закрити петлі, а малюнок вывязывается окремо. При цьому враховуйте, що кількість готових рядків, петель, які набираються на двох спицях, може різнитися зі схемою. Єдине, що потрібно — це врахування того, що готова річ вийде в розмірах менше.
Єдине, що потрібно — це врахування того, що готова річ вийде в розмірах менше.
Перший метод
Відразу визначитеся з необхідною шириною візерунка. Адже починати набирати петлі, не можна, поки не дізнаєтесь їх кількість. Кількість петельок може бути від чотирьох і більше. Якщо хочете широкий колос, то набирайте більше петель. Колоски більше підійдуть для пледів, палантинів, шарфів, а невеликі візерунки гармонують на светрах, кофтах і т. п. Ще при створенні візерунка, враховуйте товщину ниток, спиць. Коли будете набирати петлі, не забувайте про кромочні.
В’язання лицьового боку азіатського колоска спицями
Наберіть на спиці вісімнадцять петельок, далі в’яжіть таким чином:
- У першому ряду провяжите шістнадцять петельок, крім кромок, Л. П.
- У другому рядку провязывайте ці ж шістнадцять петельок виворітними.
- У третьому починайте вывязываем зернятко, розмір якого шість петельок в ширину, десять рядів у висоту.
 Схема в’язання: провяжите кромочную, 5Л.П., поворот і 6Ї.П. також в’яжіть вісім рядків.
Схема в’язання: провяжите кромочную, 5Л.П., поворот і 6Ї.П. також в’яжіть вісім рядків. - Щоб отримати новий орнамент, у тринадцятому ряду в’яжіть 6Л.П. з правої спиці, 3Л.П. з лівої спиці, після треба повернути.
- У чотирнадцятому ряду провяжите 6Ї.П., три петлі залиште без провязывания.
- Продовжуйте далі провязывают петельки, як в третьому-тринадцятому ряду. Наступний узор почнеться на 23-му ряду.
- В’яжіть полотно до необхідної довжини, в кінці у вас вийде шість петельок на спицях.
Тепер треба зв’язати іншу сторону колоса. Виконайте спочатку один ряд В. П., а другий — Л. П. Далі в’яжіть за поданою нижче схемою.
Виворітна азіатського колоска спицями:
Поверніть деталь на виворіт, продовжуйте робити петельки, але вже у зворотний бік. Якщо будете все робити по опису, то вийде ідеальний колосок з симетричним малюнком.
- У першому ряду провяжите кромочную петельку, далі 5І.П., поверніть і провяжите 6Л.
 П.
П. - У другому ряду, 3-му, 4-му, 5-му, 6-му, 7-му в’яжіть за тією ж схемою.
- У восьмому провяжите 9 петельок В. П.
- У 9-му поверніть в’язання, чергуйте І. П. з Л. П.
Коли дійдете до останнього ряду, то порівняйте половинки колоска і закрийте петлі.
ВАЖЛИВО: Якщо за зразком ви хочете розрахувати кількість рядків для певної довжини, то швидше за все у вас нічого не вийде. Під вагою виріб буде розтягуватися вниз.
Другий метод в’язання азіатського колоска спицями
Як в’язати візерунок?
Покроковий майстер-клас:
- З першого по шостий ряд в’яжіть полотно лицьовій гладдю. Якщо бажаєте рельєфний візерунок, то виконайте менше рядів лицьової гладдю, наприклад — 4-5 штук.
- Усі непарні рядки в’яжіть Л. П., а парні — В. П.
- З сьомого ряду почніть закривати середні 20 петельок і поетапно закривайте їх. Для цього провяжите десяту лицьовій петелькою, одинадцяту перенесіть з лівої спиці на праву, нитка перенесіть за неї, просто перекидання 12-ю петлю і закрийте її через одинадцяту.
І з цієї ж технології виконайте закриття всіх петельок — 20-ти штук. Потім в’яжіть Л. П.
- У восьмому ряду зв’яжіть 10Л.П., 20 петель наберіть, 10І.П. разом на спицях виходить 40 петельок.
- Повторіть з першого по восьмий ряд.
Цей раппорт ви можете повторювати до тих пір, поки не отримаєте потрібну довжину виробу. Після цього колоски збирати в один візерунок. Поверніть в’язання на іншу (виворітну сторону).
Завдяки перекрученню двох смужок колосок виходить сам по собі. А третя смужка проходить через петлю 2-ой, така схема повторюється до самого низу. Якщо ви бажаєте отримати об’ємний візерунок, простягніть всі смужки тугіше.
Азіатський колосок спицями — класичний варіант
Класична схема в’язання буде зрозуміла навіть початківцям вязальщицам. Головне — знати ази в’язання на спицях.
Перед початком процесу в’язання підготуйте пряжу, спиці необхідного розміру.
В’язання візерунка — Азіатський колосокНаберіть кількість петельок, кратне шести. В цю ж кількість досчитывайте та кромочні петлі. Далі дійте за схемою:
В цю ж кількість досчитывайте та кромочні петлі. Далі дійте за схемою:
- В першому ряду, який складається з 6-ти петель, в’яжіть Л. П. Після зупиніть процес в’язання, і переверніть полотно.
- У другому ряду в’яжіть шість В. П. І так в’яжіть шість петельок до десятогого рядка.
- Одинадцятий рядок вже в’яжеться за іншою схемою, спочатку провяжите шість петельок Л. П., після зв’яжіть ще 3Л.П. Потім можна повертати полотно.
- У дванадцятому ряд в’яжіть 6Ї.П., після поверніть в’язання і так в’яжіть десять рядків з цих шести петельок.
- За такої технології провязывайте кожен раз по 10 рядків на 6-ти петельках. Беріть при цьому 3 провязанные раніше, 3, що є на лівій спиці. В’яжіть так, поки не провяжите весь рядок.
- Після зв’яжіть два рядочки спочатку лицьовій, а після виворітного гладдю.
У другій частині процесу вся схема в’язання повторюється, але початок в’язання проводите на виворіт, і замість особових провязывайте В. П. роблять це, щоб змінити напрямок.
П. роблять це, щоб змінити напрямок.
Азіатський колосок спицями: схема в’язання візерунка на спицях по колу
Після того, як з’ясувався сам процес в’язання візерунка на двох спицях, можна приступити до в’язання азіатського колоска на кругових спицях. Метою створення цього варіанту колоска є безшовна конструкція виробу. Це не так просто, але все ж можливо.
Інструкція з в’язання азіатського колоска спицями:
- Почніть процес в’язання на круглих спицях з набору петельок. Для цього наберіть шість петель, плюс кромочная. Для зразка набираємо сорок дев’ять петель, замикаємо в коло. Потім в’яжіть весь коло лицьовими.
- У другому ряду провязывайте лише з першої по четверту петельку. І перша — це кромочная, її просто знімають. В’яжіть одинадцять рядів лицьової гладдю., останній ряд в кожному разі повинен провязываться тільки Л. П.
- У наступному ряду провяжите три петлі зі спиці в лівій руці. Зробіть поворот і в’яжіть 6 петельок. Одна петелька залишається на правій спиці — це кромочная.

- В’яжіть одинадцять рядів по 6 петельок, після введіть три петлі з лівої сторони. Продовжуйте знову на шести петлях вив’язувати одинадцять рядків лицьовій гладдю, останній ряд завжди Л. П.
- Тепер повторіть процес в’язання по всій ширині полотна. Коли вже зв’яжіть самі останні одинадцять рядків, то з’єднайте полотно в кільце. Для цього в’яжіть кромочную петельку з останньою.
- Поверніть деталь на іншу сторону — провяжите три І. П. Після чого в’яжіть тільки ці три петельки. І вывязывайте їх в особових рядах, потім останню, кромочную.
Якщо діяти за схемою, то вийде шов, але на готовій речі або аксесуар його не буде помітно. Після виконайте два ряди лицьовими петельками. Поверніть виріб, продовжуйте вив’язувати візерунок, що описаний вище в інструкції, але з вивороту. Починайте з чотирьох петельок, після прив’язуйте три петельки через одинадцять рядків кожен раз.
Коли зробите весь процес, вийде ось така деталь, як на малюнку нижче.
Після наведених описів можливо у вас залишаться якісь питання, на які дасть вам відповідь відео в кінці статті. Там наочно видно, як в’язати візерунок у вигляді азіатського колоска.
Відео: Як пов’язати азіатський колосок?
Линия 9 Баяна | Конюшня Табун д Тарасово
Родоначальник линии жеребец Баян I (Сокол-Буря) рожден в 1950 г. в колхозе «Рассвет» Островецкого района Гродненской области. Это очень крупный жеребец массивного телосложения густого упряжного типа с промерами 156-162-194-22 и живым весом 610 кг.
На ВСХВ (1957, 1958 гг.) и Всесоюзных соревнованиях в Пскове ( 1963 г.) Баян I был признан чемпионом породной группы и оценен аттестатом I степени. За 18 лет племенного использования от него получено свыше 375 голов потомства, их них 199 жеребцов и 176 кобыл. Продолжателем линии стал его лучший сын 68 Милчик, 1959 г. р.-массивная лошадь тяжелого упряжного типа. В 1965 и 1968 гг. в Пскове, в 1966 г. на ВДНХ СССР, а в 1971 г. на первой республиканской выставке в Минске Милчик оценен аттестатами I степени.
р.-массивная лошадь тяжелого упряжного типа. В 1965 и 1968 гг. в Пскове, в 1966 г. на ВДНХ СССР, а в 1971 г. на первой республиканской выставке в Минске Милчик оценен аттестатами I степени.
В 80-е годы прошлого века в линии продуцировали следующие очень ценные жеребцы, клички которых ещё можно встретить в родословных, это: Буревестник (Баян I-Березовка), Малюнок(Милчик-Мара)-аттестат I степени на ВДНХ СССР, Юпитер II (Баян III- Буланка), Полонез(Милчик-Птичка)- аттестат I степени на ВДНХ СССР, Моховик (Милчик-Машка), Плывун(Милчик-Паста), Набор (Милчик-Найда), Орган (Каштан-Сватья), Медок (Сокол-Мышка), Колосок(Милчик-Кукла), Ковер (Маяк-Кама), Разлив (102 Буян-Ромашка), Квадрат (Полонез-Капля).
На 1.01.1997 г. в линии было выделено 28 жеребцов-производителей и 63 кобылы.
На 1.01.2009 г. в планах подборов на племенных конефермах РБ значится всего 6 жеребцов-производителей и 30 конематок, самым крупным является жеребец-производитель СПК «Полесская нива « Столинского района по кличке Плут.
Действующие жеребцы-производители линии Баяна I
| № | кличка | масть | г.р. | в/х | к/д | о/г | о/п | отец | мать | хозяйство |
| 1 | Гордый 12 | гн | 2004 | 156 | 160 | 190 | 20 | Луг 16 | Горка 35 | «Судниковский» Воложинский р-н |
| 2 | Метеор 16 | гн | 2004 | 154 | 165 | 188 | 22 | Луг 16 | Машка | «Большие Новоселки» Борисовский р-н |
| 3 | Шмель 19 | мыш | 2004 | 161 | 166 | 195 | 22 | Луг 16 | Шутка 54 | «Заречье» Смолевичский р-н |
| 4 | Плут 83 | бул | 2001 | 163 | 178 | 210 | 23 | Луг 16 | Пластика | «Полесская Нива» Столинский р-н |
| 5 | Эластичный 7/31 | т. рыж рыж | 1997 | 159 | 166 | 190 | 22,5 | Спец 44 | Элита | «Достоево» Ивановский р-н |
| 6 | Баянчик | бул | 2002 | 156 | 165 | 194 | 22 | Лимон | Бандура | «Раковцы» Сморгонский р-н |
| В среднем | 158,2 | 166,7 | 194,5 | 21,9 | ||||||
| Требования класса элита | 154 | 160 | 187 | 21,5 |
В целом лошади линии Баяна I крупны, массивны и в большинстве своем тяжелого или среднего упряжного типа, широкотелы, иногда встречаются лошади с мягкой спиной, мягкими бабками. В племенной работе с этой линией следует учитывать, что наиболее удачное потомство получается при скрещивании жеребцов линии Баяна I с кобылами тяжелого типа из линий Анода I и Орлика I.
Реферат сюжетне малювання
ПланПлан
Значення та завдання сюжетного малювання.
Види і форми сюжетного малювання.
Підготовча робота до сюжетного малювання.
методика навчання сюжетного малювання у вікових групах.
Список використаної літератури.
1. ЗНАЧЕННЯ ТА ЗАВДАННЯ СЮЖЕТНОГО МАЛЮВАННЯ
У процесі сюжетного малювання діти пізнають навколишній світ. Приділяючи велику увагу малюванню, видатний радянський педагог В. О. Сухомлинський писав: «…дитячий малюнок, процес малювання — це частка духовного життя дитини. Діти не просто переносять на папір щось з навколишнього світу, а живуть у цьому світі, входять у нього як творці краси, дістаючи насолоду від неї».
Щоб виконати малюнок на якусь тему, діти повинні втілити в нього знання, нагромаджені під час цілеспрямова них екскурсій в природу і спостережень у повсякденному житті. Дошкільник, як і художник, у процесі малювання на теми іде від задуму, який опирається на його досвід.
Отже, першим етапом у творчості дитини е виникнення за-думу — теми на основі сприймання навколишньої дійсності. Щоб здійснити творчий задум, дитині треба вибрати епі-зод для малювання, уявити образи і зобразити їх на па-пері. Усе це потребує від неї мислительних процесів, уміння застосовувати знання, нагромаджені під час спостережень. Під керівництвом вихователя діти вчаться аналізувати і зображувати предмети, розміщувати їх на аркуші паперу так, щоб було зрозуміло, яку подію чи пору року зобра-жено на малюнку.
Процес зображення знайомих предметів дає можливість дітям глибше вивчити, проаналізувати, зрозуміти різно-манітність їх будови, форми, величини, пропорцій, кольору.
Під час малювання на конкретну тему розв’язуються два взаємопов’язані завдання: розвивається зорове спри-ймання в дитини і виробляється уміння осмислювати баче-не. Під розвитком сприймання ми розуміємо формування у дошкільників уміння цілеспрямовано спостерігати і по-рівнювати між собою предмети і явища, класифікувати предмети за формою і фактурою.
Знання, уміння і навички, здобуті на заняттях із сюжетного малювання, сприяють розвитку розумових здібностей дітей, уміння правильно сприймати оточуючу дійсність.
У сюжетному малюнку дитина творчо відображає все те, що справило на неї враження, збудило якісь почуття. Враження виникають у процесі безпосередніх контактів з навколишньою дійсністю, під час прослуховування роз-повідей, казок, віршів, розглядання художніх картин тощо. Ці враження створюють сприятливі умови для формування особи дитини відповідно до завдань комуністичного вихо-вання.
Сюжетне малювання в дитячому садку — могутній за-сіб патріотичного та інтернаціонального виховання дітей. Зображуючи на сюжетних малюнках радянські свята (святкування річниці Великого Жовтня, Дня Радянської Армії, 8 Березня, 1 Травня, Дня Перемоги), діти ознайом-люються з героїчним минулим і сьогоденням нашого на-роду. Це сприяє формуванню почуття гордості за свій народ.
Відображуючи на своїх малюнках будівництво сіл і міст, трудову діяльність радянських людей, успіхи радян-ської науки і техніки, діти переймаються почуттям поваги до людей праці, у них виховується любов до рідної Бать-ківщини.
У процесі виконання сюжетних малюнків у дітей вихо-вуються і певні вольові якості. Вони навчаються доводити задумане до кінця, працювати зосереджено і спрямовано, допомагати товаришеві долати труднощі, добиватися бажа-ного результату.
Сюжетне малювання дає великі можливості для есте-тичного виховання дошкільників. На заняттях та в повсяк-денному житті у дітей розвиваються естетичний смак, вміння бачити і відчувати, сприймати красу навколишньої дійсності, милуватися нею. Особливе місце в сюжетних малюнках дітей займає природа. Спостереження краси природи вихователь повинен організовувати цікаво. Видат-ний російський критик В. Г. Бєлінський писав, що необ-хідно одухотворяти для дітей весь світ і всю природу, за-ставляти говорити мовою любові й життя і німий камінь, і польову билину, і дзюркотливий струмочок, і вітерець, що тихо віє, і пурхаючого по квітах метелика…
Сюжетне малювання привчає дітей бачити багатство і різноманітність кольорів, ліній і форм оточуючих пред-метів і явищ, сприяє вихованню в них естетичних відчуттів, розвитку просторового зображення.
Діти дізнаються, що головне в сюжеті виділяється кольором і розміром, а будь-яка дія відбувається на фоні відповідного оточення. Наприклад, з народної казки «Ли-сиця і Журавель» дитина вибирає епізод і визначає кіль-кість персонажів і місце, де відбувається дія, малює го-ловних персонажів (виділяє їх розміром і кольором), а по-тім — другорядних.
Сюжетне малювання сприяє виробленню в дитини орга-нізованості, уміння обдумувати і планувати послідовність виконання малюнка, удосконалення технічних прийомів малювання олівцями і фарбами, які будуть потрібні в школі.
Таким чином, на заняттях із сюжетного малювання здійснюється розумове, моральне, естетичне та фізичне (розвиток кисті руки, її гнучкості, спритності) виховання дітей.
У «Програмі виховання і навчання в дитячому садку» (1986 р.) так визначено завдання сюжетного малювання:
1) здійснювати всебічний розвиток дітей, підготовляю-чи їх до шкільного навчання;
2) добитись розуміння ними основних вимог до компо-зиції сюжетного малюнка та ілюстрації (виявлення голов-ного в сюжеті, додержання рівноваги, ритму, формату ма-люнка і розміру зображення, відповідність кольору реаль-ності, змісту малюнка;
3) вчити дітей передавати в сюжетному малюнку фор-му, пропорції, просторове розміщення предметів, виявляти найбільш типові їх ознаки та колір; удосконалювати тех-ніку малювання кольоровим, графітним, вугільним олів-цями, гуашевими і акварельними фарбами, пастеллю;
4) розвивати творчі здібності дітей, естетичне спри-ймання навколишнього світу, просторове мислення, образну уяву, ініціативу, самостійність, зорову пам’ять, допитли-вість, інтерес до малювання, наполегливість, художній смак.![]()
Поставлені завдання реалізуються на всіх заняттях із зображувальної діяльності (малювання, ліплення, апліка-ції) та під час художньої самостійної діяльності.
У навчанні сюжетного малювання вихователь звертає особливу увагу на вміння задумувати майбутній малюнок. Н. П. Сакуліна в монографії «Малювання в дошкільному дитинстві» пише, що дитячою творчістю вихователь пови-нен керувати, що в малюванні творчість проявляється в двох основних напрямах — у формуванні зображуваль-ного задуму і передачі його в малюнку. Потрібно пам’ята-ти, що в дітей у
В Укрінформі стартував проект «Українські казки»
Укрінформ спільно з волонтерським проектом «Українська казка» та народним театром «Вавилон» започатковують проект «Українські казки».
Презентація проекту відбулася в Art Hub інформаційної агенції Укрінформ.
Про історію створення проекту Укрінформу розповіла його організатор Юлія Докторова.
«Я викладач малювання і одного разу запропонувала дітям намалювати українську казку. Я запитала, які казки вони знають, і була шокована, бо діти відповіли, що знають “Червону шапочку” та “Кіт у чоботях”. І я запропонувала таку ідею, щоб ми читали на уроках казки і за ними малювали малюнки. Разом з Укрінформом ми продовжили це як спільний проект. Укрінформ готує озвучку українських казок, розміщує їх на своєму сайті, вони будуть доповнюватися малюнками дітей”, — розповіла Докторова.
Я запитала, які казки вони знають, і була шокована, бо діти відповіли, що знають “Червону шапочку” та “Кіт у чоботях”. І я запропонувала таку ідею, щоб ми читали на уроках казки і за ними малювали малюнки. Разом з Укрінформом ми продовжили це як спільний проект. Укрінформ готує озвучку українських казок, розміщує їх на своєму сайті, вони будуть доповнюватися малюнками дітей”, — розповіла Докторова.
Казки озвучили актори київського народного театру «Вавилон». Загалом планується створити 27 подкастів.
«Наш театр долучився до цього проекту, ми намагаємося озвучувати казки так, щоб це було близько до дитячого сприйняття. Зараз у відкритому доступі є дуже багато казок, в тому числі і регіональних, наприклад, поліська казка. Ми відібрали ті, що найбільш поширені, щоб вони були максимально наближені до кожного слухача», — розповів актор театру та редактор Укрінформу Анатолій Сірик.
Відкриття виставки та презентація художньо-мистецького проекту “Українська казка” / Фото: Тарасов Володимир, УкрінформАктори озвучили вже шість казок: “Колобок”, “Колосок”, “Хитрий півник”, “Сірко”, “Півник та двоє мишенят, “Як кіт та лис ходили чоботи купувати». Вони вже доступні в розділі Подкасти на сайті Укрінформу та на YouTube-каналі агенції.
Вони вже доступні в розділі Подкасти на сайті Укрінформу та на YouTube-каналі агенції.
Також в Art Hub протягом двох тижнів триватиме виставка дитячих малюнків, які створили діти віком від 4 до 14 років за мотивами українських казок.
Організатори планують розвивати проект і надалі. Як розповів співорганізатор проекту, журналіст Укрінформу Олександр Козловський, діти можуть прослухати казки на сайті Укрінформу і намалювати свою версію казки, яка сподобалася найбільше, та надіслати малюнок до агенції.
«Це чудова нагода для дітей дати волю своєму уявленню і краще познайомитися з українськими народними казками. Це важливо, оскільки саме в дитячому віці формується світогляд, національна ідентичність. Найкращі малюнки ми будемо виставляти на сайті разом з українськими казками», — розповів Олександр Козловський.
Надіслати малюнок можна на адресу: [email protected]
Про символіку Міністерства … | від 06.10.2021 № 505/2021
Указ |
Про символіку Міністерства з питань стратегічних галузей промисловості України
1. Заснувати емблему і прапор Міністерства з питань стратегічних галузей промисловості України.
Заснувати емблему і прапор Міністерства з питань стратегічних галузей промисловості України.
2. Затвердити такі, що додаються:
Опис емблеми Міністерства з питань стратегічних галузей промисловості України;
Малюнок емблеми Міністерства з питань стратегічних галузей промисловості України;
Опис прапора Міністерства з питань стратегічних галузей промисловості України;
Малюнок прапора Міністерства з питань стратегічних галузей промисловості України;
Порядок використання емблеми та прапора Міністерства з питань стратегічних галузей промисловості України.
3. Цей Указ набирає чинності з дня його опублікування.
Президент України | В.ЗЕЛЕНСЬКИЙ |
м. Київ |
ЗАТВЕРДЖЕНО |
ОПИС
емблеми Міністерства з питань стратегічних галузей промисловості України
Емблемою Міністерства з питань стратегічних галузей промисловості України є зображення золотого птаха Фенікса із золотими перначем у правій лапі і шаблею у лівій. У центрі емблеми — малий Державний Герб України.
У центрі емблеми — малий Державний Герб України.
Емблему вміщено на темно-синій круглий щит із золотою облямівкою.
Допускається використання емблеми без щита.
Керівник Офісу |
|
ЗАТВЕРДЖЕНО |
МАЛЮНОК
емблеми Міністерства з питань стратегічних галузей промисловості України
Керівник Офісу |
|
ЗАТВЕРДЖЕНО |
ОПИС
прапора Міністерства з питань стратегічних галузей промисловості України
Прапором Міністерства з питань стратегічних галузей промисловості України є прямокутне полотнище темно-синього кольору зі співвідношенням ширини до довжини 2:3.![]()
У верхній лівій чверті прапора розміщено зображення двох рівновеликих горизонтальних смуг синього та жовтого кольорів, у центрі вільного поля полотнища — емблему Міністерства з питань стратегічних галузей промисловості України (без щита). Висота емблеми становить 1/2 ширини полотнища.
Обидві сторони полотнища прапора дзеркально ідентичні.
Керівник Офісу |
|
ЗАТВЕРДЖЕНО |
МАЛЮНОК
прапора Міністерства з питань стратегічних галузей промисловості України
Керівник Офісу | А.ЄРМАК |
ЗАТВЕРДЖЕНО |
ПОРЯДОК
використання емблеми та прапора Міністерства з питань стратегічних галузей промисловості України
1.![]() Емблема та прапор Міністерства з питань стратегічних галузей промисловості України є офіційними відмітними символами Міністерства з питань стратегічних галузей промисловості України (далі — Мінстратегпром України), що вказують на належність до цього Міністерства.
Емблема та прапор Міністерства з питань стратегічних галузей промисловості України є офіційними відмітними символами Міністерства з питань стратегічних галузей промисловості України (далі — Мінстратегпром України), що вказують на належність до цього Міністерства.
2. Емблема Мінстратегпрому України розміщується у службовому кабінеті Міністра з питань стратегічних галузей промисловості України, у залі засідань Мінстратегпрому України, використовується на прапорі Мінстратегпрому України, може відтворюватися на відомчих заохочувальних відзнаках, транспортних засобах, а також адміністративних будинках і спорудах, де розміщується Мінстратегпром України.
3. Прапор Мінстратегпрому України встановлюється у службовому кабінеті Міністра з питань стратегічних галузей промисловості України, у залі засідань Мінстратегпрому України, може встановлюватися на адміністративних будинках і спорудах, де розміщується Мінстратегпром України, використовуватися під час проведення офіційних заходів та урочистих церемоній за участю Мінстратегпрому України.
4. Відтворення зображень емблеми та прапора Мінстратегпрому України може бути виконано в одноколірному варіанті і допускається в розмірах, що відповідають меті їх застосування, із додержанням встановлених пропорцій.
5. Зображення емблеми і прапора Мінстратегпрому України може відтворюватися на друкованій рекламно-інформаційній та сувенірній продукції, у кіно-, відео- і фотоматеріалах, що видаються (виготовляються) Мінстратегпромом України або виготовляються на його замовлення.
6. В інших випадках порядок використання емблеми та прапора Мінстратегпрому України визначається Міністром з питань стратегічних галузей промисловості України.
Керівник Офісу |
|
Дом | Синцветы Stapleton (1997) и другие интерпретировали колосок (т. С-1. Один или несколько почковидных прицветников, стягивающих собственно колосок: 0 = отсутствует; 1 = присутствует, почки развиваются впоследствии или нет. С-2. Свисающий прицветник у основания оси, несущей колосок или собственно колосок: 0 = отсутствует; 1 = присутствует. С-3. Морфология выступающего прицветника: 0 = чешуйчатый или присутствует в виде рубца или ободка, не более нескольких миллиметров в длину, лезвие отсутствует; 1 = хорошо развиты как ножны, так и лезвие (часто видоизменяются). С-4. Профилл в основании оси, несущей колосок или собственно колосок: 0 = отсутствует; 1 = присутствует. С-5. Профили: 0 = целые; 1 = по крайней мере какая-то глубокая расщелина, которая разделяется вдоль на две половины. С-6. Один или несколько покрывальных прицветников, связанных с скоплениями колосков или собственно колосков: 0 = отсутствуют; 1 = присутствует. Продолжительность жизни и корневища |
 е. маленький колосок) как представляющий основной тип соцветия в семействе злаковых; эта интерпретация принята здесь. Это означает, что скопление колосков становится сложной структурой, которую мы называем синфлоресценцией.Синфлоресценции у трав традиционно описываются как колоски, кисти или метелки или некоторые вариации на одну из этих тем. Однако следует отметить, что в дополнение к сложному характеру соцветий трав они созревают по определенной схеме, так что травы, включая бамбук, не имеют настоящих колосков, кистей или метелок, за исключением самих колосков, которые прицветниковые шипы. У бамбука также существует дополнительная проблема интерпретации псевдоколоска, который содержит почки (почковидные) прицветники, которые обычно продолжают развивать дополнительные отряды псевдоколосков, но цимозным образом.Вывод гомологии среди синфлоресценций бамбука должен ждать более глубокого понимания эволюции бамбука, полученного из других источников данных, поэтому признаки в этом разделе сосредоточены в первую очередь на структурах, которые различают колоски и псевдоколоски.
е. маленький колосок) как представляющий основной тип соцветия в семействе злаковых; эта интерпретация принята здесь. Это означает, что скопление колосков становится сложной структурой, которую мы называем синфлоресценцией.Синфлоресценции у трав традиционно описываются как колоски, кисти или метелки или некоторые вариации на одну из этих тем. Однако следует отметить, что в дополнение к сложному характеру соцветий трав они созревают по определенной схеме, так что травы, включая бамбук, не имеют настоящих колосков, кистей или метелок, за исключением самих колосков, которые прицветниковые шипы. У бамбука также существует дополнительная проблема интерпретации псевдоколоска, который содержит почки (почковидные) прицветники, которые обычно продолжают развивать дополнительные отряды псевдоколосков, но цимозным образом.Вывод гомологии среди синфлоресценций бамбука должен ждать более глубокого понимания эволюции бамбука, полученного из других источников данных, поэтому признаки в этом разделе сосредоточены в первую очередь на структурах, которые различают колоски и псевдоколоски.
Вариация колоска у Zea Mays L.
 на JSTOR
на JSTOR Ботанический сад Миссури был основан в 1859 году успешным англичанином.
бизнесмена Генри Шоу и является старейшим ботаническим садом в США.На его 79 акрах расположены впечатляющие выставочные и демонстрационные сады, а также редкие
коллекции ботанических, садоводческих и исторических материалов, а также
архитектурно и исторически значимые здания. Сегодня Сад является
крупное культурное учреждение, признанное национальным историческим памятником, и
всемирно известный ботанический исследовательский центр.
Издательство ботанического сада Миссури, неотъемлемая часть исследований Сада.
подразделение, имеет более чем 100-летнюю историю научных публикаций.С
после публикации в 1890 г. статьи в «Трелизе» программа феноменально расширилась.
Включает в себя два ежеквартальных научных журнала, серию книг по различным
аспекты ботанических исследований и несколько флор, которые охватывают ботанически значимые
области мира. Анналы ботанического сада Миссури,
Основной научный журнал Гардена был основан в 1914 году и посвящен
систематическая ботаника и систематика. «Анналы» известны своим традиционным
монографии и обзоры родов растений, а также передовые статьи по
молекулярная филогенетика и ее тематические вопросы по таким темам, как происхождение
современных наземных экосистем и недавних открытий растений, животных,
и другие королевства.Новон, журнал ботанической номенклатуры, начал
в 1991 г. и содержит документы, устанавливающие новую номенклатуру сосудистых растений.
и мохообразные. Количество страниц продолжает расти из года в год,
показатель того, что важная ниша в сообществе ботанической систематики заполнена
этим журналом. Монографии по систематической ботанике Ботанического института штата Миссури.
Garden были основаны в 1978 году как способ сделать доступными длинные ботанические работы.
как хорошо сделанные, отдельные тома. Серия включает в себя несколько однотематических
трактаты, а также текущие названия, такие как Моховая флора Центральной Америки,
Icones Pleurothallidinarum, Индекс хромосомных чисел растений,
и многое другое.
«Анналы» известны своим традиционным
монографии и обзоры родов растений, а также передовые статьи по
молекулярная филогенетика и ее тематические вопросы по таким темам, как происхождение
современных наземных экосистем и недавних открытий растений, животных,
и другие королевства.Новон, журнал ботанической номенклатуры, начал
в 1991 г. и содержит документы, устанавливающие новую номенклатуру сосудистых растений.
и мохообразные. Количество страниц продолжает расти из года в год,
показатель того, что важная ниша в сообществе ботанической систематики заполнена
этим журналом. Монографии по систематической ботанике Ботанического института штата Миссури.
Garden были основаны в 1978 году как способ сделать доступными длинные ботанические работы.
как хорошо сделанные, отдельные тома. Серия включает в себя несколько однотематических
трактаты, а также текущие названия, такие как Моховая флора Центральной Америки,
Icones Pleurothallidinarum, Индекс хромосомных чисел растений,
и многое другое. MBG Press также гордится тем, что является издателем нескольких региональных и
национальные флоры, в том числе Китая, Мезоамерики, Никарагуа, Венесуэлы
Гуаяна, Панама и др. Для получения полного списка и получения дополнительной информации об исследованиях
в Ботаническом саду Миссури, см. www.mobot.org.
MBG Press также гордится тем, что является издателем нескольких региональных и
национальные флоры, в том числе Китая, Мезоамерики, Никарагуа, Венесуэлы
Гуаяна, Панама и др. Для получения полного списка и получения дополнительной информации об исследованиях
в Ботаническом саду Миссури, см. www.mobot.org.
Контроль судьбы меристем колосков кукурузы с помощью APETALA2-подобного гена indeterminate spikelet1
Abstract
Упорядоченное производство меристем с определенной судьбой имеет решающее значение для правильной разработки архитектуры растений.Меристема соцветия кукурузы несколько раз ветвится, образуя боковые меристемы с детерминированной судьбой. Первая образовавшаяся меристема, меристема пары колосков, дает две меристемы колоска, каждая из которых дает две меристемы цветка. Мы идентифицировали ген, названный индетерминантным колоском1 ( ids1 ), который определяет детерминированную судьбу меристемы колоска и тем самым ограничивает количество продуцируемых цветочных меристем. В отсутствие функции гена ids1 меристема колоска становится индетерминантной и образует дополнительные цветки.Члены семейства злаковых различаются по количеству соцветий в колоске, что позволяет предположить, что ids1 могут играть роль в архитектуре соцветий у других видов злаков. ids1 является членом семейства генов транскрипционных факторов APETALA2 ( AP2 ), которые участвуют в широком диапазоне ролей в развитии растений. Экспрессия ids1 обнаружена во многих типах зачатков латеральных органов, а также в меристемах колосков. Наш анализ мутантного фенотипа ids1 и паттерна экспрессии показывает, что ids1 специфицирует детерминированные судьбы путем подавления неопределенного роста в меристеме колосков.
В отсутствие функции гена ids1 меристема колоска становится индетерминантной и образует дополнительные цветки.Члены семейства злаковых различаются по количеству соцветий в колоске, что позволяет предположить, что ids1 могут играть роль в архитектуре соцветий у других видов злаков. ids1 является членом семейства генов транскрипционных факторов APETALA2 ( AP2 ), которые участвуют в широком диапазоне ролей в развитии растений. Экспрессия ids1 обнаружена во многих типах зачатков латеральных органов, а также в меристемах колосков. Наш анализ мутантного фенотипа ids1 и паттерна экспрессии показывает, что ids1 специфицирует детерминированные судьбы путем подавления неопределенного роста в меристеме колосков.
Ключевые слова: Меристема, колосок, колосок индетерминантный1 , APETALA2 , детерминантность
Развитие тела растения зависит от активности апикальных меристем, групп индетерминантных клеток, расположенных на верхушках растущих верхушек.![]() Способность апикальных меристем побегов оставаться неопределенной позволяет им постоянно инициировать органы, ткани и вторичные меристемы, необходимые для нормального развития. Это свойство требует, чтобы меристема выделяла популяцию клеток для своего обновления и пополнения клеток, потерянных для каждого латерального органа или вторичной меристемы.Невыполнение этого процесса приводит к прекращению роста кончика. Некоторые меристемы, такие как цветочные меристемы, потребляются при образовании боковых органов, и их можно охарактеризовать как имеющие детерминированную судьбу. Соцветия кукурузы особенно полезны для изучения детерминированности меристем, поскольку они претерпевают несколько четко определенных событий ветвления, в результате чего образуются меристемы со все более ограниченной судьбой.
Способность апикальных меристем побегов оставаться неопределенной позволяет им постоянно инициировать органы, ткани и вторичные меристемы, необходимые для нормального развития. Это свойство требует, чтобы меристема выделяла популяцию клеток для своего обновления и пополнения клеток, потерянных для каждого латерального органа или вторичной меристемы.Невыполнение этого процесса приводит к прекращению роста кончика. Некоторые меристемы, такие как цветочные меристемы, потребляются при образовании боковых органов, и их можно охарактеризовать как имеющие детерминированную судьбу. Соцветия кукурузы особенно полезны для изучения детерминированности меристем, поскольку они претерпевают несколько четко определенных событий ветвления, в результате чего образуются меристемы со все более ограниченной судьбой.
Соцветия травы организованы в единицы, называемые колоски, каждая из которых состоит из пары стерильных прицветников, называемых колосками, заключающих в себе фиксированное количество цветков (Clifford 1987). Регулирование количества цветков на колосок является основным фактором, определяющим архитектуру колосков у представителей семейства травянистых. Колосок кукурузы детерминантный, дает только два цветка в определенных положениях на оси, называемой рахиллой (Weatherwax, 1923). Родственники кукурузы характеризуются колосками с неопределенным числом цветков, как у пшеницы, или содержат только один цветочек в колоске, как у ячменя. Один классический критерий, используемый для различения различных видов трав, состоит в том, содержат ли их колоски определенное или неопределенное количество цветков (Clifford and Watson, 1977).Сам колосок является лишь одним из компонентов сложного разветвленного соцветия кукурузы. В отличие от двудольных растений, таких как Arabidopsis , у которых цветки формируются непосредственно верхушечной и латеральной меристемами (Хемпель и Фельдман, 1994), у кукурузы есть по крайней мере два отдельных этапа ветвления соцветия, прежде чем меристема колоска завершается образованием двух цветочки.
Регулирование количества цветков на колосок является основным фактором, определяющим архитектуру колосков у представителей семейства травянистых. Колосок кукурузы детерминантный, дает только два цветка в определенных положениях на оси, называемой рахиллой (Weatherwax, 1923). Родственники кукурузы характеризуются колосками с неопределенным числом цветков, как у пшеницы, или содержат только один цветочек в колоске, как у ячменя. Один классический критерий, используемый для различения различных видов трав, состоит в том, содержат ли их колоски определенное или неопределенное количество цветков (Clifford and Watson, 1977).Сам колосок является лишь одним из компонентов сложного разветвленного соцветия кукурузы. В отличие от двудольных растений, таких как Arabidopsis , у которых цветки формируются непосредственно верхушечной и латеральной меристемами (Хемпель и Фельдман, 1994), у кукурузы есть по крайней мере два отдельных этапа ветвления соцветия, прежде чем меристема колоска завершается образованием двух цветочки. Эти дополнительные ступени ветвления обеспечивают большее морфологическое разнообразие трав.
Эти дополнительные ступени ветвления обеспечивают большее морфологическое разнообразие трав.
У кукурузы был описан ряд мутантов, которые приводят к увеличению количества цветков в колоске (Veit et al.1993). У разветвленных бесшелковых мутантов дополнительные соцветия закладываются в мужских колосках метелок (Kempton 1934), хотя более экстремальная трансформация наблюдается в женских колосках колоса, в которых цветки превращаются в длинные недетерминантные ветви (Veit et al. 1993). . Изучение мутации Tasselseed6 показало, что меристема колоска претерпевает задержку в приобретении детерминированности, что позволяет ей инициировать цветки в течение более длительного периода времени (Irish et al. 1994). Анализ мутантов Tasselseed6 привел к модели (Irish 1997), в которой меристема соцветия и производные ее ветвей проходят через упорядоченный, определенный ряд детерминированных состояний развития, заканчивающийся преобразованием меристемы терминального колоска в верхний цветочек. Подобные ветвящиеся мутанты также были описаны у других видов трав и включают многоцветковую мутацию ячменя (Bossinger et al. 1992) и доминантный мутант Naked овса (Ougham et al. 1996).
Подобные ветвящиеся мутанты также были описаны у других видов трав и включают многоцветковую мутацию ячменя (Bossinger et al. 1992) и доминантный мутант Naked овса (Ougham et al. 1996).
Многочисленные генетические и молекулярные исследования выявили несколько генов, важных для развития цветков. Один такой ген, APETALA2 ( AP2 ) гомеозисного гена Arabidopsis, , выполняет несколько функций в развитии цветков, семян и семяпочек (Kunst et al.1989 год; Джофуку и др. 1994 год; Модрусан и др. 1994). Помимо своей роли в определении идентичности цветочных органов, AP2 влияет на регуляцию идентичности цветочных меристем. Например, двойные мутанты слабого аллеля ap2-1 с тождественными мутантами цветочной меристемы, такими как leafy или apetala1 , производят больше боковых ветвей кофлоресценции вместо цветков (Bowman et al. 1993). Кроме того, слабые аллели ap2 при коротком дне вызывают образование третичных цветочных побегов в пазухах трансформированных чашелистиков (Schultz, Haughn, 1993). Ген AP2 принадлежит к большому семейству генов, 12 из которых были идентифицированы у Arabidopsis (Okamuro et al. 1997). Многочисленные гомологи были идентифицированы как у однодольных, так и у двудольных (Jofuku et al., 1994; Ohme-Takagi and Shinshi, 1995; Moose and Sisco, 1997). Мутации в AP2 -подобном гене, AINTEGUMENTA, дефектны в развитии яйцеклеток (Elliot et al. 1996; Klucher et al. 1996). Недавно было показано, что глянцевый15 ген кукурузы является AP2 -подобным геном, который функционирует для подавления признаков взрослых листьев в ювенильных листьях (Moose and Sisco 1997).
Ген AP2 принадлежит к большому семейству генов, 12 из которых были идентифицированы у Arabidopsis (Okamuro et al. 1997). Многочисленные гомологи были идентифицированы как у однодольных, так и у двудольных (Jofuku et al., 1994; Ohme-Takagi and Shinshi, 1995; Moose and Sisco, 1997). Мутации в AP2 -подобном гене, AINTEGUMENTA, дефектны в развитии яйцеклеток (Elliot et al. 1996; Klucher et al. 1996). Недавно было показано, что глянцевый15 ген кукурузы является AP2 -подобным геном, который функционирует для подавления признаков взрослых листьев в ювенильных листьях (Moose and Sisco 1997).
Здесь мы описываем новый ветвящийся мутант кукурузы, индетерминантный колосок1 ( ids1 ), полученный нами при анализе фенотипа, обусловленного потерей функции кукурузы АР2 -подобного гена. Мутанты ids1 имеют индетерминантный колосок, в котором образуется несколько цветков вместо двух, встречающихся у кукурузы дикого типа. Экспрессия ids1 наблюдалась в различных боковых органах, а также в паре колосков и меристемах колосков.Наш анализ показывает, что ген ids1 имеет решающее значение для регуляции детерминированности меристемы колосков у кукурузы.
Экспрессия ids1 наблюдалась в различных боковых органах, а также в паре колосков и меристемах колосков.Наш анализ показывает, что ген ids1 имеет решающее значение для регуляции детерминированности меристемы колосков у кукурузы.
Результаты
Выделение гена ids1
Ген AP2 из Arabidopsis использовали для скрининга двух библиотек кДНК кукурузы при низкой строгости, одну из которых получали из незрелых початков, а другую из вегетативных меристем. Один и тот же класс кДНК был выделен из каждой библиотеки. Самый длинный клон этого класса состоял из 1967 нуклеотидов и содержал ORF из 433 аминокислот (рис.) с несколькими доменами, демонстрирующими поразительное аминокислотное сходство с геном AP2 Arabidopsis (Jofuku et al. 1994). Были обнаружены два тандемно повторяющихся 68-аминокислотных мотива, которые имеют 86% аминокислотную идентичность с доменом AP2 белка Arabidopsis AP2. Семейство генов AP2 можно разделить на две группы, обозначенные как EREBP-подобные или AP2-подобные, на основании наличия у них одного или двух повторов AP2 соответственно (Okamuro et al. 1997). Белок IDS1 принадлежит к последнему классу, который включает белки AP2, AINTEGUMENTA, GLOSSY15 и RAP2.7 (рис. ) (Jofuku et al., 1994; Klucher et al., 1996; Moose and Sisco, 1997; Okamuro et al., 1997). . Табачные белки ERE-BP, имеющие только один из этих повторов, связываются с ДНК (Ohme-Takagi and Shinshi 1995). Таким образом, по аналогии вполне вероятно, что IDS1 функционирует как фактор транскрипции. В подтверждение этого короткий участок основных аминокислот, который может функционировать как домен ядерной локализации, присутствует в белке IDS между аминокислотами 100 и 110 (рис.). Белок AP2 из Arabidopsis содержит богатый серином кислый домен на N-конце, который может функционировать как домен активации (Jofuku et al. 1994). Хотя аналогичная область обнаружена на амино-конце белка IDS1 в положениях 9–45, эта область намного меньше и имеет меньше кислотных и сериновых остатков по сравнению с AP2. За пределами домена AP2 существует очень небольшое сходство последовательностей между AP2 и IDS1.
1997). Белок IDS1 принадлежит к последнему классу, который включает белки AP2, AINTEGUMENTA, GLOSSY15 и RAP2.7 (рис. ) (Jofuku et al., 1994; Klucher et al., 1996; Moose and Sisco, 1997; Okamuro et al., 1997). . Табачные белки ERE-BP, имеющие только один из этих повторов, связываются с ДНК (Ohme-Takagi and Shinshi 1995). Таким образом, по аналогии вполне вероятно, что IDS1 функционирует как фактор транскрипции. В подтверждение этого короткий участок основных аминокислот, который может функционировать как домен ядерной локализации, присутствует в белке IDS между аминокислотами 100 и 110 (рис.). Белок AP2 из Arabidopsis содержит богатый серином кислый домен на N-конце, который может функционировать как домен активации (Jofuku et al. 1994). Хотя аналогичная область обнаружена на амино-конце белка IDS1 в положениях 9–45, эта область намного меньше и имеет меньше кислотных и сериновых остатков по сравнению с AP2. За пределами домена AP2 существует очень небольшое сходство последовательностей между AP2 и IDS1. Учитывая это открытие, удивительно, что положения шести интронов в домене AP2 сохраняются между ids1 и AP2 (рис.). Аналогичный результат был получен для гена gl15 кукурузы (Moose and Sisco 1997).
Учитывая это открытие, удивительно, что положения шести интронов в домене AP2 сохраняются между ids1 и AP2 (рис.). Аналогичный результат был получен для гена gl15 кукурузы (Moose and Sisco 1997).
Нуклеотидная и выведенная аминокислотная последовательность IDS1. Номера аминокислот показаны слева от ; нуклеотидов пронумерованы справа . Подчеркнутые аминокислотные последовательности представляют два домена AP2 (Jofuku et al. 1994). Серины и кислые аминокислоты на амино-конце обозначены короткими подчеркиваниями. Аминокислотные последовательности, выделенные жирным шрифтом, представляют основную область с предполагаемой активностью ядерной локализации.Нуклеотидные последовательности, выделенные жирным шрифтом, соответствуют праймеру ZAP2-1, используемому для ПЦР-скрининга. (▿) Шесть положений интрона сохраняются между IDS, AP2, и GL15; (▾) другой интрон.
Сравнение доменов AP2 AP2 -подобных генов. Выведенные аминокислотные последовательности доменов AP2 IDS1 (инвентарный номер GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AF048900″,»term_id»:»2944039″}}AF048900 ), RAP2.7 ({«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AF003100″,»term_id»:»2281638″}}AF003100), GL15 ({«type»:» entrez-нуклеотид»,»attrs»:{«текст»:»U41466″,»term_id»:»1732030″}}U41466 и ANT ({«type»:»entrez-нуклеотид»,»attrs»:{» text»:»U40256″,»term_id»:»1209098″}}U40256) сравниваются с AP2 ({«type»:»entrez-protein»,»attrs»:{«text»:»ATU12546″,»term_id» :»1275389773″}}ATU12546).Область между двумя повторами АР2, т. е. линкерная область, начинается с аминокислоты 78 и заканчивается аминокислотой 102. Промежутки, представленные точками, были введены для облегчения выравнивания. Аминокислоты, общие для всех пяти белков, выделены жирным шрифтом, а в консенсусной последовательности внизу .
Выведенные аминокислотные последовательности доменов AP2 IDS1 (инвентарный номер GenBank {«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AF048900″,»term_id»:»2944039″}}AF048900 ), RAP2.7 ({«type»:»entrez-нуклеотид»,»attrs»:{«text»:»AF003100″,»term_id»:»2281638″}}AF003100), GL15 ({«type»:» entrez-нуклеотид»,»attrs»:{«текст»:»U41466″,»term_id»:»1732030″}}U41466 и ANT ({«type»:»entrez-нуклеотид»,»attrs»:{» text»:»U40256″,»term_id»:»1209098″}}U40256) сравниваются с AP2 ({«type»:»entrez-protein»,»attrs»:{«text»:»ATU12546″,»term_id» :»1275389773″}}ATU12546).Область между двумя повторами АР2, т. е. линкерная область, начинается с аминокислоты 78 и заканчивается аминокислотой 102. Промежутки, представленные точками, были введены для облегчения выравнивания. Аминокислоты, общие для всех пяти белков, выделены жирным шрифтом, а в консенсусной последовательности внизу .
ids1 был картирован на длинном плече первой хромосомы в положении 192 с использованием рекомбинантных инбредных линий (Burr et al. 1988), зондированных неповторяющимся фрагментом ДНК с 3′-конца ids1. Никакие известные морфологические мутанты не были картированы в этой позиции.
1988), зондированных неповторяющимся фрагментом ДНК с 3′-конца ids1. Никакие известные морфологические мутанты не были картированы в этой позиции.
Выделение рецессивных мутаций ids1
Чтобы выяснить функцию ids1, , мы использовали обратную генетическую стратегию для получения аллелей потери функции. ПЦР использовали на большой популяции растений, несущих транспозоны Mutator ( Mu ), чтобы найти вставки в ids1 (Bensen et al. 1995; Meeley and Briggs 1995). Праймеры, фланкирующие домен AP2 ids1 , в сочетании с праймерами Mu давали в этом скрининге два независимых события вставки.Секвенирование продуктов ПЦР более точно локализовало вставки. Как показано на рисунке, вставка Mu в ids1-mum1 была локализована в пятом консервативном интроне домена AP2 , тогда как вставка ids1-mum2 была локализована в первом консервативном интроне на расстоянии 6 п.н. акцепторный сайт сплайсинга. Поскольку элементов Mu генерируют дупликацию 9 п.н. при вставке (Bennetzen et al. 1993), акцепторный сайт сплайсинга также обнаруживается на 5′-конце Mu.
Поскольку элементов Mu генерируют дупликацию 9 п.н. при вставке (Bennetzen et al. 1993), акцепторный сайт сплайсинга также обнаруживается на 5′-конце Mu.
Локализация элементов Mu в ids1–mum1 и ids1–mum2. Последовательности интрона и экзона ids1 показаны обычным шрифтом и жирным шрифтом соответственно. Номера интронов и экзонов показаны над последовательностями ДНК. Подчеркнутые последовательности представляют характерную дупликацию 9 п.н., связанную со вставками Mu .
Линии, несущие аллели ids1–mum1 и ids1–mum2 , демонстрировали рецессивный цветочный фенотип, более выраженный у линий ids1–mum2 .Чтобы убедиться, что вставки в ids1 ответственны за цветочный фенотип, мы проанализировали ДНК из 104 растений F2, используя ген ids1 в качестве зонда на ДНК-гелевых блотах. Косегрегация новых RFLP с цветочным фенотипом наблюдалась для 29 хромосом, содержащих аллель ids1–mum1 , и 91 хромосомы, содержащей ids1–mum2 (данные не показаны). Никаких рекомбинантов между цветочным фенотипом и ids1-mum -специфическим полиморфизмом не было обнаружено ни для одного из аллелей.
Никаких рекомбинантов между цветочным фенотипом и ids1-mum -специфическим полиморфизмом не было обнаружено ни для одного из аллелей.
Мы проанализировали фенотипы растений ids1–mum после обратного скрещивания с A632 и самоопыления. Наиболее устойчивым фенотипом у мутантов ids1–mum был дефект колоска. Зрелые колоски-метелки дикого типа содержат по два цветка, каждый из которых состоит из трех тычинок и двух лодыжек, заключенных в прицветники палеи и чешуи (рис. Б). Палея отличается от леммы своим положением и тем фактом, что она двухкилевая с двойной средней жилкой (Клиффорд и Уотсон, 1977).Колоски метелок ids1–mum2 крупнее, чем у дикого типа (рис. А), из-за наличия дополнительных цветков (рис. С). Количество цветков колеблется от не менее 3 у растений ids1–mum1 до 10 у растений ids1–mum2 . В дополнение к этим соцветиям между двумя верхними соцветиями обычно можно увидеть тонкую рахиллу, содержащую несколько более мелких цветков (рис. F). Эти более мелкие соцветия, как правило, постепенно уменьшаются, так что оставшиеся на кончике рахиллы едва видны.Более крупные дополнительные соцветия демонстрируют нормальный образец половой дифференциации и имеют правильное количество тычинок и лодыжек, окруженных лепестками и цветковыми чешуями (рис. C).
F). Эти более мелкие соцветия, как правило, постепенно уменьшаются, так что оставшиеся на кончике рахиллы едва видны.Более крупные дополнительные соцветия демонстрируют нормальный образец половой дифференциации и имеют правильное количество тычинок и лодыжек, окруженных лепестками и цветковыми чешуями (рис. C).
Фенотипы у ids1–мама мужских и женских колосков. ( A ) Обычный колосок-кисточка A632 ( левый ) и ids1–mum2 колосок-кисточка ( правый ). ( B ) Рассеченный нормальный колосок-метелка A632, показывающий боковые органы верхних и нижних цветков. ( C ) Рассеченный ids1–mum2 колосок кисточки с четырьмя цветочками вместо двух.В каждом цветке по три тычинки, окруженные цветковой чешуей и цветковой чешуей, как у А632. ( D ) Колосок нормальный А632 ( левый ) и колосок ids-mum1 ( правый ). Стрелки указывают на шелка. ( E ) Срединные продольные срезы через неоплодотворенный 25-дневный колосок ids1–mum2 ( левый ) и колосок A632 ( правый ). Черные стрелки указывают на соцветия. Колоски А632 имеют один цветочек с увеличенным нуцеллусом; ids1–mum2 колосковые колоски имеют по крайней мере три цветка с сильно редуцированным развитием боковых органов.( F ) ids1–mum2 Колосок початковый с удлиняющимся рахилом, содержащим два цветка. Единственная стрелка указывает на кончик рахиллы. (ст) Тычинка; (il) внутренняя лемма; (па) палеа; (ол) внешняя лемма; (ig) внутренняя чешуя; (og) наружная чешуя; (uf) верхний цветочек; (lf) нижний цветочек; (ра) рахилла; (фл) цветочек.
Черные стрелки указывают на соцветия. Колоски А632 имеют один цветочек с увеличенным нуцеллусом; ids1–mum2 колосковые колоски имеют по крайней мере три цветка с сильно редуцированным развитием боковых органов.( F ) ids1–mum2 Колосок початковый с удлиняющимся рахилом, содержащим два цветка. Единственная стрелка указывает на кончик рахиллы. (ст) Тычинка; (il) внутренняя лемма; (па) палеа; (ол) внешняя лемма; (ig) внутренняя чешуя; (og) наружная чешуя; (uf) верхний цветочек; (lf) нижний цветочек; (ра) рахилла; (фл) цветочек.
Женские колоски ids1–mum также имеют больший размер за счет наличия дополнительных цветков (рис. D, E). Нормальные женские колоски содержат только один зрелый цветочек из-за аборта нижнего цветка (Delong et al.1993). Развитие вторичных половых признаков у женских цветков включает аборты тычинок и развитие гинецея, о чем свидетельствует образование длинных шелков, состоящих из двух сросшихся плодолистиков (Randolph, 1936). ids1–mum1 женских колосков часто содержат более одной шелковицы, расположенной в эктопических позициях (рис. D). Однако эти шелка обычно не развиваются нормально; они часто не срослись и не могут удлиниться до нужной длины. Если развивающийся початок открыть и опылить вручную, образуется несколько зёрен, что позволяет предположить, что все остальные аспекты оплодотворения и развития плодов являются нормальными.При выращивании до зрелости эти ядра демонстрируют гомозиготных фенотипов ids1-mum и, таким образом, не представляют собой зародышевые вырезки элементов Mu . Аллель ids1–mum2 кажется более тяжелым, поскольку у большинства дополнительных цветков не развиваются полностью сформированные боковые органы (рис. E, слева). Приблизительно 5% из женских колосков ids1–mum2 имеют удлиненную рахиллу, выходящую из колоска (рис. F). В таких случаях можно заметить, что рахилла не заканчивается на самом верхнем цветке, что еще раз подтверждает ее недетерминантный характер.
ids1–mum1 женских колосков часто содержат более одной шелковицы, расположенной в эктопических позициях (рис. D). Однако эти шелка обычно не развиваются нормально; они часто не срослись и не могут удлиниться до нужной длины. Если развивающийся початок открыть и опылить вручную, образуется несколько зёрен, что позволяет предположить, что все остальные аспекты оплодотворения и развития плодов являются нормальными.При выращивании до зрелости эти ядра демонстрируют гомозиготных фенотипов ids1-mum и, таким образом, не представляют собой зародышевые вырезки элементов Mu . Аллель ids1–mum2 кажется более тяжелым, поскольку у большинства дополнительных цветков не развиваются полностью сформированные боковые органы (рис. E, слева). Приблизительно 5% из женских колосков ids1–mum2 имеют удлиненную рахиллу, выходящую из колоска (рис. F). В таких случаях можно заметить, что рахилла не заканчивается на самом верхнем цветке, что еще раз подтверждает ее недетерминантный характер. В листьях и корнях мутантов ids1 не наблюдалось очевидных и устойчивых фенотипов, несмотря на то, что ids1 экспрессируется в этих органах.
В листьях и корнях мутантов ids1 не наблюдалось очевидных и устойчивых фенотипов, несмотря на то, что ids1 экспрессируется в этих органах.
Анализ с помощью сканирующей электронной микроскопии
С помощью сканирующей электронной микроскопии определяли точку, в которой развитие меристемы колоска ids1–mum отличается от такового у дикого типа. Меристема соцветия обычно акропетально инициирует парные меристемы колосков, которые, в свою очередь, ветвятся, образуя две меристемы колосков (рис.А) (Чэн и др., 1983). Меристемы колосков также ветвятся, образуя нижние и верхние цветки. Эти соцветия начинаются в двустишном порядке, то есть чередуются на 180 градусов друг от друга (рис. B). Хотя нижняя цветочная меристема инициируется первой, развитие боковых органов происходит в верхнем цветке намного раньше, чем в нижнем (рис. C, D). Палея является первым боковым органом, который дифференцируется, и вскоре после этого развиваются три тычинки (Cheng et al. , 1983). Половая дифференциация происходит на более поздних стадиях, когда гинецей прерывается в мужских цветках, а тычинки — в женских (Dellaporta and Calderon-Urrea, 1993).
, 1983). Половая дифференциация происходит на более поздних стадиях, когда гинецей прерывается в мужских цветках, а тычинки — в женских (Dellaporta and Calderon-Urrea, 1993).
Сканирующая электронная микроскопия нормальных и ids1–mother2 ушей. ( A ) A632 зачаток колоса с инициирующими зачатками пары колосков и зачатками колосков. Пруток, 300 мкм. ( B ) Меристема неразветвленных колосков A632 ( вверху ) и чуть более старая меристема разветвленных колосков ( внизу ). Старая меристема колоска подвергается боковому разветвлению, чтобы инициировать нижний цветочек. Пруток, 102 мкм. ( C ) Развитие боковых органов в колосках A632.Первыми развиваются боковые органы верхнего цветка. Пруток, 80 мкм. ( D ) Созревающие боковые органы верхнего цветка колосковых колосков А632 с начальным гинециальным гребнем. Пруток, 104 мкм. ( E ) Ветвящиеся ids1–mum2 колосковые меристемы. Меристема самого верхнего колоска еще не разветвлена и выглядит удлиненной. Одновременное ветвление цветка и закладка чешуи происходит на противоположных сторонах средней и нижней меристемы колоска. Пруток, 156 мкм. ( F ) Дальнейшее развитие меристем колосков ids1–mum2 с удаленными колосками, показывающее развитие цветочных меристем в пазухах цветковых цветковых чешуй.Колосковая меристема сохраняется после каждого ветвления. Пруток, 58 мкм. ( G ) Old ids1–mum2 меристема колосков с удаленными колосками. Остаточная меристема колоска удлиняется, чтобы инициировать новый цветочек. Пруток, 60 мкм. ( H ) ids1–mum2 колосок с созревающими боковыми органами. Нет различий между верхним и нижним цветком с точки зрения развития боковых органов. Самый молодой цветочек, отходящий от меристемы колоска, затенен цветковой чешуей. Бар, 171 мкм.(экз/мин) меристема пары колосков; см – меристема колоска; lfm – нижняя цветочная меристема; (gy) гинецей; (fm) цветочная меристема; (ле) лемма.
Одновременное ветвление цветка и закладка чешуи происходит на противоположных сторонах средней и нижней меристемы колоска. Пруток, 156 мкм. ( F ) Дальнейшее развитие меристем колосков ids1–mum2 с удаленными колосками, показывающее развитие цветочных меристем в пазухах цветковых цветковых чешуй.Колосковая меристема сохраняется после каждого ветвления. Пруток, 58 мкм. ( G ) Old ids1–mum2 меристема колосков с удаленными колосками. Остаточная меристема колоска удлиняется, чтобы инициировать новый цветочек. Пруток, 60 мкм. ( H ) ids1–mum2 колосок с созревающими боковыми органами. Нет различий между верхним и нижним цветком с точки зрения развития боковых органов. Самый молодой цветочек, отходящий от меристемы колоска, затенен цветковой чешуей. Бар, 171 мкм.(экз/мин) меристема пары колосков; см – меристема колоска; lfm – нижняя цветочная меристема; (gy) гинецей; (fm) цветочная меристема; (ле) лемма.
Мутанты ids1–mum показывают нормальное развитие до стадии, на которой меристема колоска начинает ветвиться.![]() В этот момент меристемы колоска ids1–mum кажутся слегка удлиненными (рис. Д, вверху). В несколько более старых меристемах колосков (рис. E, в центре) можно увидеть два отдельных события ветвления, на что указывает наличие двух чешуй (стрелки).Затем в пазухах этих чешуй образуются цветочные меристемы (рис. Д, внизу и Е). Эти события ветвления происходят на противоположных сторонах меристемы колоска по дистихному образцу. Меристема колосков сохраняется после каждого ветвления и остается между двумя последними сформировавшимися цветками (рис. G, H). Эта остаточная меристема колоска продолжает инициировать дополнительные чешуйки и цветки дистихно (рис. F, G). Скорость развития боковых органов между первыми двумя цветками ids1-mum одинакова (рис.H), в отличие от дикого типа, у которого первыми развиваются органы верхнего цветка.
В этот момент меристемы колоска ids1–mum кажутся слегка удлиненными (рис. Д, вверху). В несколько более старых меристемах колосков (рис. E, в центре) можно увидеть два отдельных события ветвления, на что указывает наличие двух чешуй (стрелки).Затем в пазухах этих чешуй образуются цветочные меристемы (рис. Д, внизу и Е). Эти события ветвления происходят на противоположных сторонах меристемы колоска по дистихному образцу. Меристема колосков сохраняется после каждого ветвления и остается между двумя последними сформировавшимися цветками (рис. G, H). Эта остаточная меристема колоска продолжает инициировать дополнительные чешуйки и цветки дистихно (рис. F, G). Скорость развития боковых органов между первыми двумя цветками ids1-mum одинакова (рис.H), в отличие от дикого типа, у которого первыми развиваются органы верхнего цветка.
Мы использовали гомеобоксный ген knted1 ( kn1 ) кукурузы, который экспрессируется в меристемах, а не в детерминированных боковых органах, для отслеживания событий ветвления в колосках ids1–mum . kn1 экспрессируется в колоске и меристемах цветков кукурузы (рис. A, B) (Jackson et al. 1994). Гибридизация с антисмысловым зондом kn1 на меристемах колоска ids1–mum2 показала нормальную экспрессию в меристеме колоска (рис.С). Однако, как только меристема колоска ids1–mum2 разветвляется с образованием цветков, меристема колоска становится крупнее и, следовательно, расширяется домен экспрессии kn1 (рис. Г). Сильная экспрессия kn1 наблюдается в дополнительных цветках, начинающихся с меристемы колоска на более поздних стадиях (рис. E, стрелки), что подтверждает меристематическое происхождение дополнительных цветков. Все остальные аспекты экспрессии kn1 в цветках, включая отсутствие в боковых органах и слое L1, кажутся нормальными.
kn1 экспрессируется в колоске и меристемах цветков кукурузы (рис. A, B) (Jackson et al. 1994). Гибридизация с антисмысловым зондом kn1 на меристемах колоска ids1–mum2 показала нормальную экспрессию в меристеме колоска (рис.С). Однако, как только меристема колоска ids1–mum2 разветвляется с образованием цветков, меристема колоска становится крупнее и, следовательно, расширяется домен экспрессии kn1 (рис. Г). Сильная экспрессия kn1 наблюдается в дополнительных цветках, начинающихся с меристемы колоска на более поздних стадиях (рис. E, стрелки), что подтверждает меристематическое происхождение дополнительных цветков. Все остальные аспекты экспрессии kn1 в цветках, включая отсутствие в боковых органах и слое L1, кажутся нормальными.
Локализация in situ kn1 в меристемах колосков A632 и ids1–mum2 . ( A ) экспрессия kn1 в неразветвленной меристеме колоска A632. Пруток, 50 мкм. ( B ) kn1 экспрессия в меристеме ветвящегося колоска A632. Экспрессию можно увидеть в начальном нижнем цветочке. Пруток, 50 мкм. ( C ) kn1 экспрессия в неразветвленной меристеме ids1–mum2 колоска. Пруток, 50 мкм.( D ) kn1 экспрессия внутри меристемы раннего ветвления ids1–mum2 колоска. Инициирующие цветки можно увидеть с обеих сторон меристемы колоска. Пруток, 60 мкм. ( E ) kn1 экспрессия в более старом колоске ids1–mum2 . Сильное выражение сохраняется в развивающихся цветках. Пруток, 80 мкм. Стрелки указывают на инициирующие соцветия в B, D, и E.
( B ) kn1 экспрессия в меристеме ветвящегося колоска A632. Экспрессию можно увидеть в начальном нижнем цветочке. Пруток, 50 мкм. ( C ) kn1 экспрессия в неразветвленной меристеме ids1–mum2 колоска. Пруток, 50 мкм.( D ) kn1 экспрессия внутри меристемы раннего ветвления ids1–mum2 колоска. Инициирующие цветки можно увидеть с обеих сторон меристемы колоска. Пруток, 60 мкм. ( E ) kn1 экспрессия в более старом колоске ids1–mum2 . Сильное выражение сохраняется в развивающихся цветках. Пруток, 80 мкм. Стрелки указывают на инициирующие соцветия в B, D, и E.
Анализ экспрессии ids1
Первоначальная характеристика экспрессии ids1 с использованием РНК-гелевых блотов показала, что ген экспрессируется как в вегетативных, так и в цветочных растениях. ткани.Гель-блот РНК, зондированный 3′-частью кДНК ids1 , показывает, что транскрипт ids1 присутствует во всех тестируемых тканях (рис. A). Самая низкая степень экспрессии наблюдалась в ткани зародыша, а самая высокая — в ткани соцветия.
A). Самая низкая степень экспрессии наблюдалась в ткани зародыша, а самая высокая — в ткани соцветия.
Анализ экспрессии ids1 в различных органах кукурузы B73. ( A ) Гель-блот РНК, содержащий 1 мкг поли(A) + РНК, выделенный из эмбрионов через 17 дней после опыления ( E ), меристемы побегов, включая нераспустившийся стебель и самые молодые зачатки листьев ( M ), початок зачатки соцветий длиной до 2 см ( I ), первичные корни из семян, проросших во влажном воздухе ( R ), и пластинчатую часть полностью развернувшихся ювенильных листьев ( L ) промокли и гибридизовали с 3′ частью кДНК ids1 , которая не включает домен AP2.( B ) Контроль гибридизации. Блот в A исследовали с кДНК убиквитина кукурузы, чтобы показать относительную нагрузку.
Для определения уровней транскрипта ids1 у мутантов ids1–mum поли(А) + РНК выделяли из ушей мутантов и сравнивали с РНК из ушей дикого типа. Как показано на рисунке A, ни в одном аллеле ids1-mum не было обнаружено транскриптов дикого типа. Вместо этого наблюдались слабые транскрипты с более высокой молекулярной массой, которые могут быть результатом химерных или альтернативно сплайсированных транскриптов.Этот результат демонстрирует, что вставки элементов Mu в ids1 не подвергаются сплайсингу и что оба ids1-mum1 и ids1-mum2 могут представлять аллели потери функции.
Как показано на рисунке A, ни в одном аллеле ids1-mum не было обнаружено транскриптов дикого типа. Вместо этого наблюдались слабые транскрипты с более высокой молекулярной массой, которые могут быть результатом химерных или альтернативно сплайсированных транскриптов.Этот результат демонстрирует, что вставки элементов Mu в ids1 не подвергаются сплайсингу и что оба ids1-mum1 и ids1-mum2 могут представлять аллели потери функции.
Анализ экспрессии ids1 в мутантных ушах ids1 – mum . ( A ) Гель РНК, содержащий 1 мкг поли(А) + РНК, выделенной из 2-сантиметровых ушей A632 ( 1 ), ids1–mum2 ( 2 ) и ids1–mum1 ( 3 ) подвергали блоттингу и гибридизовали с 3′-частью кДНК ids1 .( B ) Контроль гибридизации. Блот в A исследовали с кДНК убиквитина кукурузы, чтобы показать относительную нагрузку.
Чтобы более точно локализовать временной и пространственный паттерн экспрессии ids1 , мы использовали гибридизацию in situ. На рисунке А показаны продольные срезы верхушки вегетативного побега дикого типа, зондированного с помощью меченной дигоксигенином антисмысловой РНК, полученной из 3′-конца клона ids1 . Положение листьев внутри побега кукурузы было описано с помощью индекса пластохрона (Lamoreaux et al.1978), в котором самый молодой лист обозначается как пластохрон 1 или P 1 , а последующие листья обозначаются как P 2 , P 3 и т. д. Положение зарождающегося листа в меристеме обозначается как P 0 . Экспрессия ids1 обнаруживается на флангах меристемы в дискретной зоне (рис. А). Исходя из положения самого молодого листа (обозначенного как P 1 ), мы предполагаем, что эта зона экспрессии меристемы находится в положении, в котором будет инициироваться следующий лист (обозначенный как P 0 ).Экспрессия также наблюдается на кончиках двух следующих более старых листьев, P 1 и P 2 .
На рисунке А показаны продольные срезы верхушки вегетативного побега дикого типа, зондированного с помощью меченной дигоксигенином антисмысловой РНК, полученной из 3′-конца клона ids1 . Положение листьев внутри побега кукурузы было описано с помощью индекса пластохрона (Lamoreaux et al.1978), в котором самый молодой лист обозначается как пластохрон 1 или P 1 , а последующие листья обозначаются как P 2 , P 3 и т. д. Положение зарождающегося листа в меристеме обозначается как P 0 . Экспрессия ids1 обнаруживается на флангах меристемы в дискретной зоне (рис. А). Исходя из положения самого молодого листа (обозначенного как P 1 ), мы предполагаем, что эта зона экспрессии меристемы находится в положении, в котором будет инициироваться следующий лист (обозначенный как P 0 ).Экспрессия также наблюдается на кончиках двух следующих более старых листьев, P 1 и P 2 .
Локализация in situ в вегетативных и цветочных апексах B73 с использованием 3′-части кДНК ids1 . ( A ) Верхушечная меристема побега с зачатками молодых листьев (P 1 и P 2 ) и зачатками зачатков листьев (P 0 ). Пруток, 65 мкм. ( B ) Соцветие молодого колоса. Экспрессию ids1 можно увидеть в акропетально инициирующих парных зачатках колосков.Пруток, 70 мкм. ( C ) Колосковые меристемы неразветвленные. Экспрессию можно увидеть в меристеме колосков, но не в колосках. Пруток, 36 мкм. ( D ) Колосковые меристемы ветвящиеся. Сильное выражение можно увидеть во внутренней и внешней цветковых чешуях, а также в зоне между начальными верхними и нижними цветками (немаркированная стрелка). Эта зона экспрессии постепенно исчезает в более старшем колоске внизу. В инициирующих цветочных меристемах экспрессии не обнаружено. Пруток, 70 мкм. ( E ) Верхний цветочек с начальными тычинками.Отдельные области экспрессии ids1 можно наблюдать в инициирующих зачатках тычинок и палеях. Экспрессия ids1 сохраняется в расширяющихся внутренней и внешней цветковых чешуях.
( A ) Верхушечная меристема побега с зачатками молодых листьев (P 1 и P 2 ) и зачатками зачатков листьев (P 0 ). Пруток, 65 мкм. ( B ) Соцветие молодого колоса. Экспрессию ids1 можно увидеть в акропетально инициирующих парных зачатках колосков.Пруток, 70 мкм. ( C ) Колосковые меристемы неразветвленные. Экспрессию можно увидеть в меристеме колосков, но не в колосках. Пруток, 36 мкм. ( D ) Колосковые меристемы ветвящиеся. Сильное выражение можно увидеть во внутренней и внешней цветковых чешуях, а также в зоне между начальными верхними и нижними цветками (немаркированная стрелка). Эта зона экспрессии постепенно исчезает в более старшем колоске внизу. В инициирующих цветочных меристемах экспрессии не обнаружено. Пруток, 70 мкм. ( E ) Верхний цветочек с начальными тычинками.Отдельные области экспрессии ids1 можно наблюдать в инициирующих зачатках тычинок и палеях. Экспрессия ids1 сохраняется в расширяющихся внутренней и внешней цветковых чешуях. Пруток, 70 мкм. ( F ) Старые соцветия с лодикулами. Выражение тычинок теперь локализовано в камерных камерах пыльника. В лодикулах (lo) не видно никакого выражения. Пруток, 70 мкм. ( G ) ids1–мама2 колосок, прозондированный ids1. Может быть обнаружена небольшая часть транскрипта ids1 или его отсутствие.Стрелки указывают на цветочки. Пруток, 130 мкм.
Пруток, 70 мкм. ( F ) Старые соцветия с лодикулами. Выражение тычинок теперь локализовано в камерных камерах пыльника. В лодикулах (lo) не видно никакого выражения. Пруток, 70 мкм. ( G ) ids1–мама2 колосок, прозондированный ids1. Может быть обнаружена небольшая часть транскрипта ids1 или его отсутствие.Стрелки указывают на цветочки. Пруток, 130 мкм.
В продольных срезах цветочной ткани экспрессия ids1 обнаружена в нескольких различных зачатках латеральных органов, а также в паре колосков и меристемах колосков. На рисунке B продольные срезы меристем раннего соцветия, прозондированные с помощью ids1 , показывают полосы экспрессии в повторяющемся узоре, который, очевидно, соответствует зачаткам пар колосков. На более поздней стадии, после формирования колосков на ножке и сидячих колосков, экспрессия ids1 обнаруживается по всей меристеме колоска, но не в первых двух зачатках боковых органов, образованных из меристемы колоска, наружной и внутренней чешуйках (рис. С). Вскоре после инициации цветочных меристем (рис. D) экспрессия ids1 присутствует в инициирующих внутренних и наружных листьях прицветника чешуи, стягивающих цветки, но отсутствует в самой цветочной меристеме. Кроме того, в области между верхним и нижним цветками видна сильная полоса экспрессии (рис. D, немеченая стрелка). В немного более старых колосках, как показано в нижней части рисунка D, эта полоса экспрессии начинает исчезать. Сканирующая электронная микрофотография колосков на эквивалентных стадиях показана на рисунке B.В более зрелых соцветиях кистей (рис. E) дискретные группы клеток цветочной меристемы, расположенные для формирования тычинок и палео, экспрессируют ids1. Экспрессия сохраняется на более поздних стадиях в более старых расширенных лепестках и цветковых чешуях, а также в локулярных камерах пыльника (рис. F). В лодикулах или гинецее початка наблюдалась небольшая экспрессия или ее отсутствие (рис. F; данные не показаны). Как показано на рисунке G, при использовании антисмыслового зонда ids1 на колосков ids1-mum экспрессия ids1 не обнаруживается.
С). Вскоре после инициации цветочных меристем (рис. D) экспрессия ids1 присутствует в инициирующих внутренних и наружных листьях прицветника чешуи, стягивающих цветки, но отсутствует в самой цветочной меристеме. Кроме того, в области между верхним и нижним цветками видна сильная полоса экспрессии (рис. D, немеченая стрелка). В немного более старых колосках, как показано в нижней части рисунка D, эта полоса экспрессии начинает исчезать. Сканирующая электронная микрофотография колосков на эквивалентных стадиях показана на рисунке B.В более зрелых соцветиях кистей (рис. E) дискретные группы клеток цветочной меристемы, расположенные для формирования тычинок и палео, экспрессируют ids1. Экспрессия сохраняется на более поздних стадиях в более старых расширенных лепестках и цветковых чешуях, а также в локулярных камерах пыльника (рис. F). В лодикулах или гинецее початка наблюдалась небольшая экспрессия или ее отсутствие (рис. F; данные не показаны). Как показано на рисунке G, при использовании антисмыслового зонда ids1 на колосков ids1-mum экспрессия ids1 не обнаруживается.![]()
Обсуждение
Регулирование числа цветков, инициированных меристемой колоска, является важным этапом в определении морфологии колоска. Мутации в гене ids1 влияют на детерминированность меристемы колосков. Вместо образования только верхнего и нижнего цветков меристема колосков ids1–mum продолжает инициировать дополнительные цветки в двудольной филлотаксии. Понимание того, как ids1 способствует детерминированности меристемы колосков, требует изучения различных моделей инициации цветков кукурузы.
Одна из моделей закладки цветков предполагает, что нижний цветочек закладывается латерально, тогда как верхний цветочек является конечным, образующимся в результате трансформации меристемы колоска в верхний цветочек (рис. A) (Irish 1997). У некоторых видов злаков, таких как anthoxanthum, , колосок оканчивается цветком, а семязачаток является конечной структурой (Clifford 1987). Если верхнее соцветие является терминальной структурой при нормальном развитии кукурузы, белок IDS1 может функционировать, способствуя трансформации меристемы колоска в верхнее соцветие.![]() IDS1 может делать это самостоятельно или посредством активации генов идентичности меристем цветков, таких как ортологи LEAFY и APETELA1 , которые могут трансформировать меристемы соцветий в меристемы цветков (Mandel and Yanofsky 1995; Weigel and Nilsson 1995). Если бы это было так, мы бы предположили, что трансформация меристемы колоска в меристему цветка задерживается у мутантов ids1-mum , что позволяет меристеме колоска сохраняться и инициировать большее количество цветков. Такая интерпретация использовалась для объяснения фенотипа доминантных Ts6 мутантов кукурузы, у которых формируются дополнительные соцветия (Irish 1997).Однако мутанты Ts6 отличаются от мутантов ids1–mum тем, что они также влияют на детерминацию пола и вызывают отсутствие подавления плодолистика внутри метелки, но не в ухе.
IDS1 может делать это самостоятельно или посредством активации генов идентичности меристем цветков, таких как ортологи LEAFY и APETELA1 , которые могут трансформировать меристемы соцветий в меристемы цветков (Mandel and Yanofsky 1995; Weigel and Nilsson 1995). Если бы это было так, мы бы предположили, что трансформация меристемы колоска в меристему цветка задерживается у мутантов ids1-mum , что позволяет меристеме колоска сохраняться и инициировать большее количество цветков. Такая интерпретация использовалась для объяснения фенотипа доминантных Ts6 мутантов кукурузы, у которых формируются дополнительные соцветия (Irish 1997).Однако мутанты Ts6 отличаются от мутантов ids1–mum тем, что они также влияют на детерминацию пола и вызывают отсутствие подавления плодолистика внутри метелки, но не в ухе.
Модели для закладки соцветий кукурузы. ( A ) Модель верхушечного цветка. Меристема колоска разветвляется один раз латерально, образуя нижний цветочек. Остаточная меристема колоска (показана черным) затем превращается в верхний цветочек (черный). ( B ) Модель бокового разветвления.Меристема колоска подвергается боковому разветвлению с образованием нижних, а затем верхних цветков. Остаточная меристема колосков (показана черным цветом) находится в небольшой области между цветками. Зачаток меристемы колоска обнаружен у рахиллы в зрелом колоске (черный).
Остаточная меристема колоска (показана черным) затем превращается в верхний цветочек (черный). ( B ) Модель бокового разветвления.Меристема колоска подвергается боковому разветвлению с образованием нижних, а затем верхних цветков. Остаточная меристема колосков (показана черным цветом) находится в небольшой области между цветками. Зачаток меристемы колоска обнаружен у рахиллы в зрелом колоске (черный).
Альтернативная модель предполагает, что и верхние, и нижние цветки являются латеральными продуктами меристемы колосков. Два соцветия кукурузы были интерпретированы как боковые ветви рахиллы с небольшими остатками оси между соцветиями или без них (Bonnett 1953).Если ветвление верхнего цветка боковое, то после закладки верхнего цветка в рахилле может быть обнаружен сильно редуцированный зачаток меристемы колоска (рис. Б). Согласно этой модели, IDS1 должен участвовать в подавлении индетерминантного роста внутри меристемы колоска, тем самым препятствуя регенерации меристемы колоска после ветвления верхнего цветка. У мутантов ids–mum остаточная меристема колосков пролиферирует и продолжает инициировать цветки.
У мутантов ids–mum остаточная меристема колосков пролиферирует и продолжает инициировать цветки.
На основании паттерна экспрессии ids1 и времени возникновения дефектов ids1-mum мы отдаем предпочтение второй модели и выдвигаем гипотезу, что IDS1 функционирует для подавления неопределенного роста внутри меристемы колоска.Согласно первой модели, переключение судьбы меристемы колоска происходит только после закладки нижнего цветка. Однако дефекты в меристемах колосков ids1–mum сначала наблюдались в меристеме колосков до закладки нижнего цветка (рис. Д). Время возникновения дефекта указывает на то, что изменение в детерминации судьбы происходит раньше, в соответствии с экспрессией ids1 , наблюдаемой в меристеме колоска до того, как происходит ветвление цветка (Fig. C). Первая модель также предполагает, что вся меристема колоска превращается в верхний цветочек, не оставляя остаточной меристемы колоска.Тем не менее, экспрессия ids1 была обнаружена в зоне между верхними и нижними цветками у дикого типа (рис. D), в которой, как можно было бы ожидать, располагалась остаточная меристема колосков. Дополнительным доказательством того, что эта область представляет собой остаточную меристему колосков, является наблюдение, что она регенерирует после ветвления цветков у мутантов ids1-mum (Fig. F, G). Кроме того, ожидается, что первая модель ids1 будет экспрессироваться в будущем верхнем цветке, чтобы способствовать его трансформации в цветочную идентичность.Вместо этого, как показано на рисунке D, мы не видим экспрессии ids1 ни в верхней, ни в нижней меристеме цветка. Наконец, если бы детерминированность меристемы колоска просто задерживалась у мутантов ids1-mum в соответствии с первой моделью, можно было бы ожидать, что рахилла в конце концов оканчивается цветком. Этого не наблюдалось ни у самца, ни у самки ids1–mum колосков (рис. Е, данные не представлены).
D), в которой, как можно было бы ожидать, располагалась остаточная меристема колосков. Дополнительным доказательством того, что эта область представляет собой остаточную меристему колосков, является наблюдение, что она регенерирует после ветвления цветков у мутантов ids1-mum (Fig. F, G). Кроме того, ожидается, что первая модель ids1 будет экспрессироваться в будущем верхнем цветке, чтобы способствовать его трансформации в цветочную идентичность.Вместо этого, как показано на рисунке D, мы не видим экспрессии ids1 ни в верхней, ни в нижней меристеме цветка. Наконец, если бы детерминированность меристемы колоска просто задерживалась у мутантов ids1-mum в соответствии с первой моделью, можно было бы ожидать, что рахилла в конце концов оканчивается цветком. Этого не наблюдалось ни у самца, ни у самки ids1–mum колосков (рис. Е, данные не представлены).
Считается, что развитие меристемы критического размера у кукурузы является сигналом для возникновения ветвления (Sundberg and Orr 1996). Один механизм, с помощью которого IDS1 может подавлять неопределенный рост, заключается в ограничении размера остаточной меристемы колосков, так что она больше не может инициировать цветки. У мутантов ids1-mum размер остаточной меристемы колоска может быть разрегулирован и увеличен до большего, чем обычно. Более крупная меристема колоска потенциально может позволить большее количество событий ветвления для образования дополнительных цветков. В подтверждение этой идеи меристема колоска ids1-mum выглядит более удлиненной, чем обычно, до закладки цветков (рис.Е). Кроме того, больший диаметр меристемы колосков наблюдается вскоре после инициации первого цветка у мутантов ids1-mum (рис. D).
Один механизм, с помощью которого IDS1 может подавлять неопределенный рост, заключается в ограничении размера остаточной меристемы колосков, так что она больше не может инициировать цветки. У мутантов ids1-mum размер остаточной меристемы колоска может быть разрегулирован и увеличен до большего, чем обычно. Более крупная меристема колоска потенциально может позволить большее количество событий ветвления для образования дополнительных цветков. В подтверждение этой идеи меристема колоска ids1-mum выглядит более удлиненной, чем обычно, до закладки цветков (рис.Е). Кроме того, больший диаметр меристемы колосков наблюдается вскоре после инициации первого цветка у мутантов ids1-mum (рис. D).
Однако важно признать, что судьба меристемы не всегда зависит от размера меристемы. Например, верхняя меристема цветка намного больше, чем нижняя меристема цветка во время инициации (рис. B), и тем не менее обе они в равной степени настроены на образование цветков. Таким образом, наличие более крупной меристемы колоска не обязательно наделяло бы ее большей неопределенностью.Не менее важным компонентом различия между детерминантными и индетерминантными судьбами меристем колоска является способность к регенерации после закладки цветка. Меристемы с неопределенной судьбой могут проявлять большую способность к саморегенерации независимо от их исходного размера. Измененная морфология, наблюдаемая в меристемах колосков мутантов ids1-mum , может быть просто отражением этой способности.
Таким образом, наличие более крупной меристемы колоска не обязательно наделяло бы ее большей неопределенностью.Не менее важным компонентом различия между детерминантными и индетерминантными судьбами меристем колоска является способность к регенерации после закладки цветка. Меристемы с неопределенной судьбой могут проявлять большую способность к саморегенерации независимо от их исходного размера. Измененная морфология, наблюдаемая в меристемах колосков мутантов ids1-mum , может быть просто отражением этой способности.
Возможно, что IDS1 функционирует для поддержания детерминированности меристемы колоска, репрессируя те факторы, которые необходимы для поддержания неопределенности.Гомеобоксный ген kn1 экспрессируется в некоторых типах неопределенных меристематических тканей, а не в детерминированных органах, таких как листья (Smith et al., 1992; Jackson et al., 1994). Мутанты с потерей функции kn1 в кукурузе демонстрируют уменьшенное ветвление метелок и меньшее количество колосков. Предполагается, что этот фенотип является результатом потери неопределенных клеток соцветия, которые в отсутствие kn1, приняли детерминированную судьбу (Kerstetter et al. 1997). Если kn1 на самом деле поддерживает неопределенность меристемы, то экспрессия kn1 должна сохраняться в меристеме колоска ids1-mum и показывать расширенный домен.Хотя это наблюдалось (рис. D), важно отметить, что kn1 в норме экспрессируется как в колоске, так и в меристемах цветков (рис. A, B). Т.о., тот факт, что домен экспрессии kn1 расширяется у мутантов ids1-mum , может просто отражать дополнительные области инициации цветков.
Предполагается, что этот фенотип является результатом потери неопределенных клеток соцветия, которые в отсутствие kn1, приняли детерминированную судьбу (Kerstetter et al. 1997). Если kn1 на самом деле поддерживает неопределенность меристемы, то экспрессия kn1 должна сохраняться в меристеме колоска ids1-mum и показывать расширенный домен.Хотя это наблюдалось (рис. D), важно отметить, что kn1 в норме экспрессируется как в колоске, так и в меристемах цветков (рис. A, B). Т.о., тот факт, что домен экспрессии kn1 расширяется у мутантов ids1-mum , может просто отражать дополнительные области инициации цветков.
Ген ids1 был клонирован путем гомологии с геном AP2 Arabidopsis , и было показано, что он содержит два повтора высококонсервативного домена AP2.Хотя белок IDS1 демонстрирует большее сходство с AP2 по сравнению со всеми другими AP2 -подобными генами, клонированными до сих пор, ген ids1 может не представлять собой фактический ортолог AP2 кукурузы. Аминокислотная гомология между IDS1 и AP2 не распространяется за пределы домена AP2. Более того, экспрессия ids1 не была обнаружена в плодолистиках, как это было обнаружено для AP2 (Jofuku et al. 1994). Наконец, предварительные эксперименты показывают, что сверхэкспрессия кДНК ids1 в Arabidopsis не дополняет мутантный фенотип ap2-1 (G.Чак неопубликован). Дополнительные члены семейства генов AP2 , вероятно, присутствуют в геноме кукурузы, поскольку гибридизации низкой строгости с доменом ids1 AP2 в качестве зонда на Саузерн-блотах кукурузы показывают несколько гибридизирующих полос (данные не показаны). Кроме того, было показано, что несколько тегов экспрессируемых последовательностей из библиотек кДНК кукурузы имеют домены AP2 (Klucher et al. 1996).
Аминокислотная гомология между IDS1 и AP2 не распространяется за пределы домена AP2. Более того, экспрессия ids1 не была обнаружена в плодолистиках, как это было обнаружено для AP2 (Jofuku et al. 1994). Наконец, предварительные эксперименты показывают, что сверхэкспрессия кДНК ids1 в Arabidopsis не дополняет мутантный фенотип ap2-1 (G.Чак неопубликован). Дополнительные члены семейства генов AP2 , вероятно, присутствуют в геноме кукурузы, поскольку гибридизации низкой строгости с доменом ids1 AP2 в качестве зонда на Саузерн-блотах кукурузы показывают несколько гибридизирующих полос (данные не показаны). Кроме того, было показано, что несколько тегов экспрессируемых последовательностей из библиотек кДНК кукурузы имеют домены AP2 (Klucher et al. 1996).
Даже несмотря на то, что ids1 экспрессируется в зачатках вегетативных и цветочных боковых органов, явных мутантных фенотипов в этих органах обнаружено не было. В этих органах может действовать определенный уровень генетической избыточности, выявить который можно только при анализе двойных мутантов. Например, двойные мутанты ap2 и aintegumenta демонстрируют фенотипы цветочных органов, не наблюдаемые ни у одного мутанта Arabidopsis (Elliot et al. 1996). Принимая во внимание большое количество AP2 -подобных генов, присутствующих в геноме Arabidopsis (Okamuro et al. 1997), неудивительно обнаружить такую функциональную избыточность, встречающуюся у кукурузы.Недавно было высказано предположение, что два близкородственных гена MADS box, zag1 и zmm2, , играют перекрывающиеся роли в спецификации идентичности плодолистиков и тычинок у кукурузы (Mena et al. 1996).
В этих органах может действовать определенный уровень генетической избыточности, выявить который можно только при анализе двойных мутантов. Например, двойные мутанты ap2 и aintegumenta демонстрируют фенотипы цветочных органов, не наблюдаемые ни у одного мутанта Arabidopsis (Elliot et al. 1996). Принимая во внимание большое количество AP2 -подобных генов, присутствующих в геноме Arabidopsis (Okamuro et al. 1997), неудивительно обнаружить такую функциональную избыточность, встречающуюся у кукурузы.Недавно было высказано предположение, что два близкородственных гена MADS box, zag1 и zmm2, , играют перекрывающиеся роли в спецификации идентичности плодолистиков и тычинок у кукурузы (Mena et al. 1996).
Хотя цветочные органы ids1–mum соцветий метелок полностью функциональны и напоминают органы дикого типа, дефекты цветочных органов наблюдались у ids1–mum женских колосков. Возможное объяснение этого дефекта может заключаться в том, что у нормальных женских колосков отмирает нижний цветочек. Если различие между верхними и нижними цветками утеряно у мутантов ids1–mum , то все цветки могут получать сигнал аборта, который обычно присутствует только в нижних цветках дикого типа. Для проверки этой модели будет полезен анализ двойных мутантов с мутантом tasseled 2 , у которого абортов нижних цветков не происходит (DeLong et al. 1993).
Если различие между верхними и нижними цветками утеряно у мутантов ids1–mum , то все цветки могут получать сигнал аборта, который обычно присутствует только в нижних цветках дикого типа. Для проверки этой модели будет полезен анализ двойных мутантов с мутантом tasseled 2 , у которого абортов нижних цветков не происходит (DeLong et al. 1993).
В свете того факта, что многие представители семейства злаковых обладают индетерминантными колосками, возникает соблазн предположить, что AP2 -подобные гены играют роль в регуляции количества цветков у других злаков.Предполагается, что колоски с множеством цветков являются древним признаком, учитывая, что производные виды почти повсеместно развили уменьшение количества цветков (Stebbins 1987). Возможный механизм такой редукции мог включать изменение функции или домена экспрессии гена ids1 с включением меристемы колоска в дополнение к различным латеральным органам, где он обычно экспрессируется. В этом отношении будет интересно проанализировать экспрессию ids1 как в современных, так и в древних злаках.
Структура травы
Структура травыБольшая часть этого раздела получена или основана на материале из Травы. Нового Южного Уэльса (Wheeler et al., 1982)
Вегетативные персонажи
Вегетативная или нецветущая часть травянистое растение весьма изменчиво. Растения могут расти густыми пучками или беспорядочно альпинисты. В то время как вегетативные признаки часто легко различимы, у трав они часто чрезвычайно изменчивы, легко поддаются влиянию климата и питания. условия.
Привычка
Привычка описывает основной «внешний вид» растения, который может быть довольно изменчивым. в травах. Это варьируется от символов основного размера до других, описывающих шероховатость или опушенность различных листовых частей растения. Цветущие стебли включены с вегетативными признаками, чтобы избежать путаницы, поэтому высота стебля относится к максимальной высоте растения, независимо от того, цветет оно или нет.
Продолжительность Продолжительность жизни трав непостоянна. Некоторые могут возникать только в течение короткого времени
после дождя и их циклы могут быть не привязаны к временам года (эфемерные). Другие
завершают свой жизненный цикл в течение года и начинают с семян каждый год (ежегодно).
Третьи сохраняются в течение двух (двухлетних) или более (многолетних) лет. Это может
быть трудным характером для определения. Если растение трудно выдернуть
и имеет массу корней и остатки старых побегов вокруг основания, это может
хорошо быть многолетним. Если растение относительно стройное и с тонкими корнями,
скорее всего годовой.
Некоторые могут возникать только в течение короткого времени
после дождя и их циклы могут быть не привязаны к временам года (эфемерные). Другие
завершают свой жизненный цикл в течение года и начинают с семян каждый год (ежегодно).
Третьи сохраняются в течение двух (двухлетних) или более (многолетних) лет. Это может
быть трудным характером для определения. Если растение трудно выдернуть
и имеет массу корней и остатки старых побегов вокруг основания, это может
хорошо быть многолетним. Если растение относительно стройное и с тонкими корнями,
скорее всего годовой.
Многие травы растут компактными пучками или кочками, но другие дают побеги
которые растут горизонтально и укореняются в каждом узле. Это полезно для растения, потому что
если побег сломается, он продолжит расти как отдельное растение. Если бегуны
встречаются над землей, они называются столонами и корневищами если они происходят под землей. Узлы столонов дают зеленые листья,
в то время как маленькие коричневые чешуйчатые листья образуются на узлах корневищ.
Побеги образуются из пазушных почек, расположенных под углом к стеблю. и листовое влагалище встречаются. Если новый побег растет между стеблем и листовым влагалищем он называется интравагинальным . Если он прорывается через основание листового влагалища это называется экстравагинальным побегом.
СтеблиСтебель – это вертикальный стебель в середине побега (или побега). они твердые в узлах и часто полые на всем протяжении междоузлия.В некоторых травах междоузлия стебля очень короткие, пока стебель не удлиняется для цветения. Стебли варьируются от неразветвленных (простых) до многоразветвленных.
ЛистьяЛист травы представляет собой удлиненную структуру, возникающую в узле и состоящую из базального цилиндрического влагалища , которое окружает стебель или более молодые листья. Над ним уплощенная лопасть с параллельным жилкованием.
Листовые ножны представляют собой полые цилиндры
разделите одну сторону так, чтобы поля обычно перекрывались. Ухоподобные выступы
(ушные раковины) часто образуются в верхней части листового влагалища. Ушные раковины могут быть
редуцируется до опушенного края основания листовой пластинки или может встречаться на одной стороне
только лист. Ушные раковины, хотя и являются отличительной чертой многих видов, тонкие.
структур и может увядать с возрастом. Молодые листья следует всегда осматривать при
работа с ушными раковинами.
Ухоподобные выступы
(ушные раковины) часто образуются в верхней части листового влагалища. Ушные раковины могут быть
редуцируется до опушенного края основания листовой пластинки или может встречаться на одной стороне
только лист. Ушные раковины, хотя и являются отличительной чертой многих видов, тонкие.
структур и может увядать с возрастом. Молодые листья следует всегда осматривать при
работа с ушными раковинами.
Полоска механической ткани, часто бледная, на стыке лезвия и ножен, абаксиальная часть которого называется воротником .(Абаксиальный относится к поверхность, обращенная в сторону от оси растения). Слой абсциссии может также развиваются, позволяя лезвию сбрасываться.
A язычок в форме языка или
лоскутообразная прозрачная мембрана или волоски, прикрепляется к обращенной к поверхности листа
стержень (адаксиальная поверхность) в месте соединения лезвия с оболочкой. Язычковый тип
особенно полезный идентификационный признак, поскольку он относительно постоянен
внутри рода. У некоторых трав могут отсутствовать язычки, например. Echinochloa crus-galli (Скотная трава).Многие злаки имеют полупрозрачные перепончатые язычки, в то время как
у других язычки окаймлены волосками или в виде ободка из волосков.
У некоторых трав могут отсутствовать язычки, например. Echinochloa crus-galli (Скотная трава).Многие злаки имеют полупрозрачные перепончатые язычки, в то время как
у других язычки окаймлены волосками или в виде ободка из волосков.
В основании листового влагалища и пластинки вставочных меристем что для всех листьев, кроме старых, они позволяют листу удлиняться после удаления лезвие в результате скашивания или выпаса скота или повреждения ветром и дождем. Интеркалярные меристемы области, в которых клетки сохраняют свою способность делиться и производить новые клетки. Поскольку листья травы созревают от кончика лезвия до основания, меристема оболочки остается активной дольше, чем меристема пластинки.Интеркалярные меристемы также встречаются у трав у основания междоузлий стебля, где активность позволяет стебель расти вертикально после полегания.
Форма, текстура, складчатость, опушенность и т. д. листовой пластинки могут быть полезными диагностическими признаками, они часто варьируются в пределах одного вида. или даже на том же заводе. У некоторых листовых пластинок средняя жилка выражена.
часто бледного цвета, у других нервы одинаково заметны или незаметны. Вернация часто определяется как расположение молодого листа в
бутон, здесь мы определили вернацию как расположение листа при осмотре
в поперечном разрезе (поперек листа).Листовые пластинки часто плоские, но могут
сворачиваться или складываться (скручивание или складывание является реакцией на водный стресс в некоторых
случаях), а некоторые настолько узкие, что напоминают щетину.
или даже на том же заводе. У некоторых листовых пластинок средняя жилка выражена.
часто бледного цвета, у других нервы одинаково заметны или незаметны. Вернация часто определяется как расположение молодого листа в
бутон, здесь мы определили вернацию как расположение листа при осмотре
в поперечном разрезе (поперек листа).Листовые пластинки часто плоские, но могут
сворачиваться или складываться (скручивание или складывание является реакцией на водный стресс в некоторых
случаях), а некоторые настолько узкие, что напоминают щетину.
Персонажи соцветий
Основные символы соцветия Стебель травяного стебля в репродуктивной стадии оканчивается одним или несколькими соцветий или цветковых головок. Соцветие включает в себя все
структуры над последним листом стебля или основания. В некоторых тропических травах
то, что сначала выглядит как соцветие, представляет собой сложный набор ветвей
с листовидной структурой, покрывало или покрывало , у основания
каждого отдельного соцветия. Ветви над спатеолой могут быть уплотнены
или модифицированы и часто находятся парами. Покрывала могут быть цветными или зелеными и
может или не может быть дифференцирован на лезвие и ножны.
Ветви над спатеолой могут быть уплотнены
или модифицированы и часто находятся парами. Покрывала могут быть цветными или зелеными и
может или не может быть дифференцирован на лезвие и ножны.
Каждое соцветие состоит из нескольких колоски, содержащие очень мелкие цветки, расположенные на ветвях или главная ось. Колоски, единицы соцветия травы, либо прикрепляются непосредственно к ветвям, и в этом случае они сидячие , или прикрепляются с помощью стеблей или цветоножек .Стебель сразу под соцветие цветонос , который продолжается основной осью или rachis после первой ветки или колоска. «Рахис» является общим термином и часто используется для любой оси или ветви, к которой прикреплены колоски.
В этом ключе мы разделили соцветия по их основной форме при взгляде на
на расстоянии. Соцветие в форме твердого пера или эскимо.
если смотреть на расстоянии вытянутой руки, называется твердым , а соцветие с
очевидные ветви соединение соцветие.Соцветия с двумя
или более ответвлений, происходящих из одной и той же точки, являются цифрами , или если
ветви отходят почти от одной и той же точки подпальцевой .
Хотя соцветие может состоять из колосков, расположенных непосредственно на стебле, основной тип травянистого соцветия – сложная метелка , обычно называемый просто метелкой, со вторичным и часто более высоким порядком ветвления. Ветка высшего порядка может состоять из колоса с прикрепленными к нему колосками прямо, тогда как у кисти колоски прикрепляются к рахису по цветоножкам.Ветви как с сидячими, так и с черешковыми колосками технически rames , но здесь рассматриваются как кистевидные. появляются многочисленные соцветия на расстоянии быть одним стержнем на самом деле являются промежуточными по типу и могут можно рассматривать как метелки с уменьшенным ветвлением. Если ветки короткие и скученные, метелка узкая и описывается как колосовидная , суженный или плотный. Уменьшение числа ветвей может привести к первичная ось с кистями или шипами.
КолосокКаждый колосок состоит из ряда цветков или соцветий расположенных попеременно по обе стороны от центральной оси, rhachilla , выше двух пустые прицветники, колосковых чешуек . «Прицветник» — это общий термин для значительно уменьшенного лист, особенно мелкие листья, стягивающие цветки. Травяной цветок заключен двумя прицветниками, внешний из которых на стороне, удаленной от оси, называемой леммой и внутренний прицветник, названный palea .Прицветники вместе с цветком, составить цветочек.
Колоски в соцветии обычно похожи внешне, но двух типов могут встречаться в одном и том же соцветии или в разных соцветиях. В Ламаркия стерильные колоски, не дающие плодов, смешиваются с фертильными колосками.
Колоски многих родов трибы Andropogoneae рождаются разнородными парами, причем один член пары плодовит (и часто сидячие) и другие ножки, стерильные и более мелкие, они называются компаньонами . стерильные колоски .
Мужские и женские колоски могут встречаться в отдельных соцветиях, которые часто непохожий внешне. У однодомного растения, такого как Zea mays (кукуруза), оба типы соцветий встречаются на одном и том же растении. В Zea mays самец соцветие верхушечное, самка (початок) находится в пазухах нижние листья.
У двудомного растения, такого как Spinifex sericeus Мужские и женские соцветия находятся на разных растениях. У Cleistochloa колоска с цветками, которые открытые нормально для опыления (хазмогамные) встречаются в соцветиях одного типа, в то время как клейстогамные колоски (которые не раскрываются) встречаются в соцветиях, заключенных в листовых влагалищах.Колоски или соцветия, которые встречаются в двух формах, часто называют диморфным.
Многие травы, кроме упомянутых андропогоноидных типов, имеют колоски. организованы в группы, т.е. Hordeum (Ячмень).
Степень зрелости колосков, вероятно, варьирует в пределах большого соцветия. Часто более старые колоски располагаются ближе к верхушке или середине соцветия. Желательно и обычно возможно исследовать колоски на разных стадиях развития. зрелости при идентификации травы.
ОберткаКолоски или группы колосков могут быть покрыты различными структурами, которые выполняют функцию рассеивания семян. Эти структуры включают волоски, щетинки, шипы, заусенцами или стерильными ветвями и называются оберткой.
РачиллаЭта маленькая ось изогнута попеременно в противоположных направлениях (изгиб или «зигзаг»). и несет цветочки, расположенные в два ряда.
Колосковые чешуиКаждый колосок имеет две чешуи на база.Колосковые чешуи никогда не стягивают цветок или палею. Однако они могут быть изменены и различаются по размеру, форме, текстуре и т. д., и один (или редко оба) могут отсутствовать. Полоски на чешуе представляют собой тяжи сосудистой ткани (вены), называемые нервов . Количество и выраженность нервов у разных видов различаются. но достаточно постоянна внутри вида. Колосковые чешуи могут напоминать приведенные выше леммы. их и важно при осмотре травянистых соцветий уметь распознавать колосковые чешуи и, таким образом, идентифицировать колоски как единицы.
ЦветочкиСамой внешней структурой цветочка является лемма , которая обычно заключает в себе другие части. При сжатии с боков лемма обычно имеет один ребро. или киль идущий по середине спины. Обычно видны и часто постоянными по численности для определенного рода или вида, являются несколько жилок или нервы . Нижние цветковые чешуи часто зеленые и перепончатые, особенно в незрелом состоянии. и становятся более твердыми и часто менее зелеными по мере взросления.Существует значительный вариации в размере, форме, цвете, текстуре, сжатии, опушенности и наличии придатков типа ости (см. вариации). Такой вариации особенно полезны при идентификации.
У основания многих цветков (как у Aristida и Austrostipa ) представляет собой затвердевшую структуру, часто остроконечную, которая развивается по мере созревания и хорошо видно, когда цветочек отделяется от рахилы. Это мозоль может быть образован из части рахиллы, а также из части цветковой чешуи и полезен для рассеивания семян, позволяя цветковым чешуям прилипать к шкурам животных, и проникать в почву после того, как устройство было рассредоточено.Острая мозоль Hordeum murinum (трава ячменя) вызывает слепоту у ягнят и травы, такие как Aristida spp. (Проволочная трава) значительный ущерб животным прячется. Жесткое, обычно заостренное основание колоска (как у Heteropogon , Andropogon и родственные роды) сразу над точкой расчленения также называется мозолью. В данном случае это часть рахиллы. В Eriochloa мозоль представляет собой утолщенный узел и остаток первой чешуи; в Chrysopogon входит в состав цветоножки.
Paleas обычно имеют 2 киля (и 2 жилки), более мягкие и менее изменчивые по структуре, чем леммы. В соцветии плоская сторона обращена к рахилле. в то время как кили касаются сторон чешуи, охватывая, таким образом, цветок. Палеас бамбука и Oryza sativa (рис) и их родственники лишь частично заключены в их леммах.
ЦветыЧашелистиков и лепестков нет, но есть два или три небольших мясистых образования, lodicules , в основании яичника, считаются редуцированным околоцветником.Лодикулы — тонкие структуры лежащий между завязью и цветковой чешуей (и палией, если присутствует третья). В виде цветок раскрывается (антезис) они набухают, раздвигая цветковые цветковые чешуи и позволяют сначала выступить пыльникам (обычно), а затем рыльцам (проект за прицветниками), что позволяет опыление. Protandry состояние в у которых первыми появляются тычинки и протогиния при появлении рыльца перед пыльниками. Когда рыльца экспонируются в течение нескольких часов, лодикулы сморщиваются, позволяя цветковым чешуям и палеям закрыться над развивающимся зерном.
Прикрепляются к основанию цветка кольцом или мутовкой внутри лодикул, 1-6 тычинок. Большинство злаков имеют 3 тычинки с длинными тонкими нитями. Пыльники, открывающиеся щелями, универсальны; это нити крепятся в одной точке над основанием. Это позволяет пыльнику раскачиваться ветер, позволяющий пыльце разлетаться во всех направлениях.
Обычно удлиненный верхний яичник содержит один очаг или камера, с одной семяпочкой , прикрепленной у основания на бледной стороне.На вершине завязи расположены два боковых столбика (у некоторых злаков сросшиеся). для придания единого центрального стиля) с волосатыми рыльцами. В ядовитых злаках рыльца бледные, в то время как у панцирных злаков обычно темные (от розовых до черных).
Зерно Семяпочка цветка травы развивается после
оплодотворения, в семя с большим количеством крахмалистого эндосперма и зародышем, лежащим на
одна сторона. Стенка завязи, образующая околоплодник плода, в норме
сросшиеся с семенной оболочкой и плодом, называемым зерновкой или зерном . Sporobolus и Eleusine образуют семянки, в которых семенная
оболочка) свободна от околоплодника. Некоторые бамбуки дают ягоды. Большинство травяных зерен
сбрасываются, заключенные в лемму и палею. Форма, поверхность, текстура, относительная
размер и положение эмбриона являются полезными диагностическими признаками.
Многие систематики трав считают основным («типичный») рисунок колосков похож на многоцветковые колоски травы, такие как Avena sativa (овес).Считается, что это было изменено у других видов добавлением таких структур, как ости и волоски и т.д. и на уменьшение количества и/или размера частей колоска.
ДополненияВолоски, щетинки, шипы и т.п. могут быть прикреплены к частям колосков. В некоторых иногда такие структуры окружают отдельные колоски или группы колосков. Самый очевидный придаток — ость.
Ость представляет собой длинный тонкий придаток, прикрепленный к кончику, спинке или основанию колосковой чешуи, цветковой чешуи или чешуи, а место прикрепления является диагностическим признаком.Некоторые ости закручены по спирали, многие скручиваются и раскручиваются в ответ на изменения влажности и вызывающие основание цветковой чешуи (часто с резкой каллюсной мозолью) проникать в почву или прятаться. Такие ости делятся на две части, относительно толстая колонка и концевая тонкая щетина . Очень тонкая ость часто называют щетиной , а очень короткую концевую ость mucro .
СокращенияМожно уменьшить как количество, так и размер частей колоска.Номер количество цветков в колоске колеблется от одного до 20, а у некоторых злаков колоски редуцированы только до чешуи.
Если отсутствует только завязь, соцветие называют тычиночным или самец , хотя его часто также называют бесплодным или бесплодным. Цветочек без тычинки или завязи, с листовой чешуей и цветковой чешуей или только с цветковой чешуей, называется стерильным или neuter , хотя также используются термины бесплодный или пустой. цветочек с яичником описывается как плодородных .Термины стерильный, фертильный и т. могут применяться в качестве определителей к леммам соответствующих цветочков.
Хотя травянистые соцветия обычно обоеполые, однополые цветки (оба мужские и женский) не редкость. Могут встречаться в колосках с обоеполыми цветками, в однополых колосках или в соцветиях с колосками только одного пола.
Размеры колосковых, цветковых и черешковых чешуй также могут быть уменьшены. В Микролаэна stipoides (плакучая трава) колосковые чешуи редуцированы до крошечных чешуек, а у Лолиум сп.(Райграс) одна чешуйка отсутствует на всех, кроме верхушечного колоска. Стерильные цветковые чешуи некоторых Phalaris spp. может быть 1 мм в длину.
Другие вариантыТо, как распадаются колоски при созревании, может быть полезным диагностическим признаком. особенность. Сустав может присутствовать на рахилле над колосковой чешуей, что позволяет соцветия опадают по мере созревания, оставляя колосковые чешуи на цветоножке. Альтернативно колосковые чешуи могут опадать вместе с колоском, и в этом случае говорят, что колосок отделить ниже чешуи.Дезартикуляция может иметь место и в суставы рахиллы и позвоночника. Парные колоски трав, принадлежащие в трибу Andropogoneae обычно попадают как единица вместе с частью стержень кисти, к которой они прикреплены.
Рахилла продолжается за самый верхний цветочек в виде маленькой щетинки в какие-то ядовитые колоски. Это «продолжение» рахиллы, часто трудно видеть, полезно в идентификации.
Имеет значение и положение редуцированных цветков в колоске.Уменьшенные соцветия может возникать на рахиллах выше или ниже фертильных соцветий.
Некоторые из упомянутых вариаций, как правило, встречаются вместе и характеризуют две основные группы злаков. Ядовитые травы имеют от одного до многих соцветий на колосок. с редуцированными соцветиями над фертильными цветками и расчленением над колосками. Паникоидные травы демонстрируют то, что часто называют «уменьшенным» рисунком колосков. У этих злаков число цветков снижено до двух, стерильность (редуцированная) соцветие ниже фертильного соцветия на очень короткой рахилле и расчленение находится ниже чешуи.Конечно, есть много исключений из этих обобщений.
Топ
Границы | OsVIL2 регулирует развитие колосков, контролируя регуляторные гены у Oryza sativa
Введение
У злаков есть уникальная единица соцветия, колосок, который содержит разное количество цветков и колосковых чешуек в зависимости от вида (Bommert et al., 2005). Колосок риса ( Oryza sativa ) состоит из двух рудиментарных чешуй, двух пустых чешуй и одного цветка (Bommert et al., 2005). Каждый цветочек состоит из цветковой чешуи, цветка, двух лодыжек на стороне цветка, шести тычинок и плодолистика (Yoshida, Nagato, 2011).
Гены семействаAPETALA2/фактор отклика на этилен (AP2/ERF), SUPERNUMERARY BRACT ( SNB ) и INDETERMINATE SPIKELET1 ( IDS1 ) играют решающую роль в переходе от колосковой меристемы (SM) к цветочной меристеме (SM) (FM) (Ли и др., 2007; Ли и Ан, 2012). Их мутантные линии производят повторяющиеся чешуйки и демонстрируют аномальный рисунок цветочных органов из-за расширенной активности SM.Другой ген AP2/ERF, MULTI-FLORET SPIKELET1 ( MFS1 ), также играет роль в регуляции судьбы SM. У мутантов mfs1 образуются дополнительные леммоподобные органы и удлиненные рахиллы, а пустые чешуи и палеа дегенерируют (Ren et al., 2013). Эти результаты предполагают, что правильный переход от SM к FM необходим для нормального развития колосков.
Анализ различных мутантов риса выявил несколько генов, участвующих в развитии чешуи. Например, мутации в EXTRA GLUME1 ( EG1 ) и EG2 / OsJAZ1 , которые функционируют в передаче сигналов жасмоновой кислотой, вызывают аномальные фенотипы колосков.Пустые чешуйки трансформируются в чешуевидные органы и образуются дополнительные чешуйки (Li et al., 2009; Cai et al., 2014). Кроме того, затрагиваются идентичность и количество цветочных органов. Эти изменения, вероятно, связаны с измененной экспрессией OsMADS1 у мутантов (Jeon et al., 2000a; Prasada et al., 2005). Сходные фенотипы наблюдаются для растений риса INDETERMINATE GAMETOPHYTE1 ( OsIG1 ) RNAi (Zhang et al., 2015). Мутации в длинной стерильной лемме ( G1 ) связаны с гомеотической трансформацией стерильной леммы в лемму, что позволяет предположить, что ген репрессирует идентичность леммы, чтобы специфицировать стерильную лемму (Yoshida et al., 2009). Мутации OsMADS34 вызывают плейотропные эффекты, включая превращение пустых чешуй в чешуевидные органы (Gao et al., 2010).
Развитие palea задерживается у мутантов, дефектных по RETARDED PALEA1 ( REP1 ) (Yuan et al., 2009). DEPRESSED PALEA 1 ( DP1 ), кодирующий ДНК-связывающий белок AT-hook, также играет решающую роль в развитии palea (Jin et al., 2011). Мутации этого гена вызывают дефект палеа, а также увеличение количества цветочных органов.Анализ экспрессии показал, что DP1 функционирует выше по течению от REP1 . Мутации OsMADS15 также приводят к дефектной палео (Wang et al., 2010), в то время как мутации OsMADS6 также вызывают нарушение палео и изменение развития плодолистика (Ohmori et al., 2009; Li et al., 2010). Мутации OsMADS32 связаны с дефектными маргинальными областями для palea и эктопических органов цветка (Sang et al., 2012).
Белки группыPolycomb (PcG) являются эпигенетическими репрессорами, контролирующими экспрессию генов (Mozgova, Hennig, 2015).Они функционируют в различных процессах развития, образуя репрессивный комплекс 2 Polycomb (PRC2), который ингибирует хроматин-мишень посредством триметилирования лизина 27 гистона 3 (h4K27me3) (Cao et al., 2002; Czermin et al., 2002; Müller et al. , 2002; Некрасов и др., 2005). PRC2 контролирует инициацию FM, спецификацию идентичности органов и терминацию меристемы (Gan et al., 2013).
PRC2 состоит из нескольких компонентов. У Arabidopsis мутанты основных компонентов PRC2 — CLF/SWN, FIE, EMF2 и MSI — представляют собой эктопическую экспрессию AGAMOUS ( AG ), вызывая аномальные цветочные фенотипы, сходные с фенотипами AG -сверхэкспрессии. растения (Гудрич и др., 1997; Йошида и др., 2001; Хенниг и др., 2003 г.; Мун и др., 2003; Кац и др., 2004). Этот комплекс также влияет на терминацию FM, регулируя временную экспрессию WUSCHEL ( WUS ) и KNUCKLES ( KNU ) (Мозгова и др., 2015). После инициации всех цветочных органов FM прекращается посредством репрессии WUS , гена поддержания меристемы (Mayer et al., 1998). PRC2 ингибирует экспрессию KNU , который ингибирует транскрипцию WUS (Lenhard et al., 2001; Сан и др., 2009). Для терминации цветков активированный AG вытесняет PRC2 из KNU , что приводит к активации KNU и репрессии WUS (Sun et al., 2014).
VERNALIZATION INSENSITIVE 3 (VIN3), другой компонент PRC2, усиливает h4K27me3 в FLOWERING LOCUS C ( FLC ), репрессоре цветения (Sung and Amasino, 2004). У риса VIN3-подобный белок OsVIL2 усиливает цветение, опосредуя h4K27me3 на хроматине LFL1 , который является негативным регулятором цветения (Yang et al., 2013). OsVIL2 связывается с OsEMF2b, ортологом Arabidopsis EMF2 (Yang et al., 2013). Мутанты OsEMF2b демонстрируют фенотипы серьезных дефектов цветочных органов и неопределенность меристемы, сходные с мутантами, дефектными по генам E-функции OsMADS1, OsMADS6 и OsMADS34 (Luo et al., 2009; Yang et al., 201 ; Conrad et al., 2014; Xie et al., 2015). OsEMF2b подавляет экспрессию этих генов E-функции, изменяя h4K27me3 в их хроматине (Conrad et al., 2014). Кроме того, OsEMF2b ингибирует OsLFL1 и OsMADS4 путем опосредования h4K27me3 на их хроматине, что приводит к стимулированию цветения и регулированию спецификации идентичности цветочных органов (Xie et al., 2015).
Мутанты osvil2 также демонстрируют аномальное развитие колосков. В этом исследовании мы изучили мутантные фенотипы, которые появляются на ранних стадиях этого процесса, и провели анализ транскриптома для идентификации нижестоящих генов, контролируемых OsVIL2 .
Материалы и методы
Растительные материалы и условия роста
Две мутантные линии OsVIL2 , osvil2-1 и osvil2-2 , были выделены из пула меченых линий Т-ДНК риса (Jeon et al., 2000b; Jeong et al., 2002; Yang et al. ., 2013). Семена мутантов и дикого типа (WT) проращивали на среде MS и проводили генотипирование для идентификации гомозиготных растений, как объяснялось ранее (Yi and An, 2013). Растения выращивали либо на рисовом поле в естественных условиях, либо в теплице при дополнительном искусственном освещении.
Векторное строительство и преобразование риса
Для конструкции слияния промотора OsVIL2 и GUS мы использовали фрагмент промотора длиной 2348 п.н. между -2340 и +8 из сайта инициации трансляции OsVIL2 , который использовали для комплементации мутанта (Zhao et al., 2010). Фрагмент амплифицировали с помощью ПЦР и помещали выше гена GUS без промотора с использованием бинарного вектора pGA3519, который содержит селектируемый маркер гигромицина.Последовательности праймеров для амплификации промоторной области перечислены в дополнительной таблице S1. Agrobacterium tumefaciens , штамм LBA4404, трансформировали этим вектором методом замораживания-оттаивания (An et al., 1989). Трансгенные растения, экспрессирующие репортерный ген GUS , получали совместным культивированием клеток Agrobacterium с щитковыми каллюсами, полученными из зрелых семян риса (сорт Longjin). Совместно культивируемые каллусы отбирали и регенерировали, как сообщалось ранее (Jeon et al., 1999).
Гистохимическое окрашивание GUS
Ткани растений погружали в раствор для окрашивания GUS, содержащий 100 мМ фосфата натрия (рН 7,0), 0,1 мМ феррицианида калия, 0,1 мМ ферроцианида калия, 0,1% тритона Х-100, 10 мМ ЭДТА (рН 8,0), 1% ДМСО, 0,1% X-gluc (5-бром-4-хлор-3-индолил-β- D -глюкуроновая кислота/циклогексиламмониевая соль) и 5% метанол (Yoon et al., 2014). Образцы инкубировали в течение ночи при 37°C, затем переносили в 70%-ный этанол при 65°C на несколько часов для удаления хлорофиллов перед хранением в 95%-ном этаноле.
Сканирующая электронная микроскопия
Образцы готовили, как описано ранее (Lee et al., 2007). Образцы фиксировали в растворе FAA, обезвоживали в серии этанола и сушили в критической точке в Leica EM CPD300 (Leica Microsystems, Wetzlar, Germany). Их устанавливали на заглушки, напыляли платиной и наблюдали под сканирующим электронным микроскопом (SIGMA FE-SEM, Carl Zeiss, Германия).
Выделение РНК и анализ qRT-PCR
ВсегоРНК выделяли с помощью RNAiso Plus (TaKaRa, Shiga, Япония).кДНК синтезировали с использованием 2 мкг РНК, обратной транскриптазы вируса мышиного лейкоза Молони (Promega, Мэдисон, Висконсин, США), ингибитора РНКазина рибонуклеазы (Promega), 10 нг олиго (dT) 18 праймера и 2,5 мМ дезоксирибонуклеотидтрифосфаты. Количественную ОТ-ПЦР проводили с использованием SYBR Green I Prime Q-Mastermix (GENETBIO, Тэджон, Южная Корея) и Rotor-Gene 6000 (Corbett Research, Сидней, Новый Южный Уэльс, Австралия) в соответствии с протоколами, о которых сообщалось ранее (Yang et al., 2013). ; Вэй и др., 2016). Рис Ubiquitin1 использовали в качестве внутреннего контроля для количественной ПЦР в реальном времени (qRT-PCR).
Для анализа транскриптома тотальные РНК получали из 2- и 4-мм метелок растений WT и osvil2-1 в трех биологических повторностях. Двухцепочечные кДНК синтезировали со случайными гексамерами и лигировали с адаптерами. Эта библиотека была секвенирована парами концов с использованием стратегии PE90 на Illumina HiSeq TM 2000 в Пекинском институте геномики (Ухань, Китай).Алгоритм DEseq был применен для фильтрации дифференциально экспрессируемых генов. Все аннотации Gene Ontology (GO) были загружены с веб-сайта NCBI, UniProt и Gene Ontology. Данные секвенирования РНК депонированы в базу данных GEO (номер доступа: GSE108538).
Результаты
Мутации в
OsVIL2 вызвали аномальное образование колосковРанее мы показали, что линии osvil2-1 и osvil2-2 с мутантными инсерциями Т-ДНК демонстрируют плейотропные фенотипы, включая позднее цветение, меньшее количество побегов и аномальное развитие колосков (Yang et al., 2013). В этом исследовании мы подробно охарактеризовали дефекты колоска (рисунок 1 и дополнительный рисунок S1). Колоски WT имели пару рудиментарных чешуй плюс пустые чешуи (рис. 1А). Напротив, количество рудиментарных колосковых чешуй было увеличено у 14% колосков osvil2 (рис. 2А). Кроме того, пустые чешуи были удлинены в 21% мутантных колосков (рис. 1B и дополнительная фигура S1B), в то время как количество пустых чешуй уменьшилось на 21%, в результате чего появились колоски с одной пустой чешуей или без нее (рис. 1C, 2B). .Иногда на месте пустой чешуи развивался чешуевидный орган. Эти наблюдения показали, что мутации OsVIL2 влияли как на количество, так и на морфологию пустых чешуек.
РИСУНОК 1. Колосковые фенотипы WT и мутант osvil 2. (A–E) Фенотипы WT и osvil2-1 колоски. (А) Колосок WT с парой рудиментарных чешуй и пустыми чешуями, нижняя цветковая чешуя, палея. (Б) освил2-1 колосок с удлиненными пустыми чешуями. (C) osvil2-1 колосок с вырожденной палеей и без пустой чешуи со стороны цветковой чешуи (острие стрелки). (D) osvil2-1 колосок с дополнительным чешуевидным органом. (E) osvil2-1 колосок с 2 цветковыми чешуями и 2 вырожденными палеями. (F–K) Фенотипы внутренних цветочных органов WT и osvil2-1 . Палеа и лемма были удалены. (F) Колосок WT состоит из двух лодыжек на стороне листовой чешуи, шести тычинок (звездочки) и одного пестика с двумя рыльцами (стрелки). (G) osvil2-1 цветочек с дополнительными лодикулами и незрелыми тычинками. (H) osvil2-1 цветочек с лодикулярно-тычиночными мозаичными органами. (I) osvil2-1 цветочек с дополнительными лодикулами, семью тычинками (звездочками) и двумя плодолистиками. (J) освил2-1 плодолистик, в котором срослось несколько плодолистиков. Также увеличено количество клейм (стрелки). (K) Лодикуло-тычиночный мозаичный орган в osvil2-1 . (L) Поперечный срез колоска WT. (М,Н) Сечение Освил2 Колосок. (М) Образование дополнительного чешуевидного органа у osvil2 колоска. (N) osvil2 с двумя нижними цветковыми чешуями, двумя вырожденными палеями, тремя аномальными лодикулами (остриями стрелок) и одним сросшимся плодолистиком с двумя семязачатками. боп, тело палеи; Са, плодолистик; ДП, депрессивная бледная кожа; EG, пустая чешуя; ЭЭГ — удлиненная пустая чешуя; L — лемма; Лодикула; L/S, лодикуло-тычиночный мозаичный орган; Мне нравится; леммоподобный орган; mrp — краевая область палео; Ов, семязачаток; Р, палеа; RG, рудиментарная чешуя.Звездочками обозначены тычинки (F,I) или сосудистые пучки (L–N) . Масштабные линейки = 1 мм (A–K) или 500 мкм (L–N) .
РИСУНОК 2. Номера органов цветка в WT, osvil2-1 и osvil2-2 колосков. (A) Рудиментарные чешуи; (B) пустые чешуи; (C) лемма и палея; (D) лодикулы; (Е) тычинки; (F) пестик.
Кроме того, у osvil2 соцветий было аномальное развитие всех органов цветка.Дополнительные подобные лемме структуры наблюдались в 45% колосков osvil2 , создавая три или более таких структур (рис. 1D, 2C и дополнительный рисунок S1C). Дополнительные чешуевидные органы часто образовывались на эктопическом обороте. Некоторые из этих эктопических леммоподобных органов напоминали лемму (рис. 1D). Число цветковых чешуй было увеличено в 14% колосков из osvil2 , и они часто сопровождали вырожденные палеа (рис. 1Д, 2В). Развитие palea было дефектным у 33% этих мутантов (рис. 1C и дополнительный рисунок S1D), а поперечные срезы колосков показали дополнительные леммоподобные органы (рис. 1M) и дегенерированные palea (рис. 1N).
Количество лодикул увеличилось до трех и более в 54% колосков osvil2 (рис. 1G-I, 2D и дополнительные рисунки S1F, G), и их морфология иногда была аномальной. Лодикулы были удлиненными (20%) или в верхней части 26% этих лодикул образовался пыльникоподобный орган (рис. 1 З, I, К). Число тычинок уменьшилось у 8% колосков (рис. 1З, 2Д), но увеличилось у 22% (рис. 1И, 2Д). Наконец, количество плодолистиков возросло до двух у 32% мутантных колосков (рис. 1I, 2F), и они часто были сросшимися (рис. 1 H, J).Эти наблюдения показали, что OsVIL2 необходим для правильного развития всех органов внутри колоска.
Нарушено раннее развитие колосков у
osvil2Мы использовали сканирующий электронный микроскоп для изучения дефектов колосков на ранних стадиях развития, которые были классифицированы на основе предыдущих исследований (Ikeda et al., 2004). В WT FM продуцировал пустую чешуйку и чешуйку на противоположной стороне пустой чешуи во время стадии развития колоска Sp3 в WT (рис. 3A).Палеа впоследствии развилась на стадиях 4–5 (рис. 3В). В то время как нижняя цветковая чешуя и цветная чешуя удлинялись, тычинки развивались на стадии 6 (рис. 3С). Наконец, в центре колоска на стадии 8 возник плодолистик (рис. 3D).
РИСУНОК 3. Сканирование электронных микрофотографий колосков WT и osvil2 на ранних стадиях развития. (A–D) WT колоски. (А) Стадия Sp3 при формировании зачатков цветковых чешуй. (Б) Стадия Sp4-5 формирования палео зачатков; возможно, внутри чешуи закладываются два зачатка лодикулы. (В) Стадия Sp6, с образованием шести тычиночных зачатков (звездочка). (D) Стадия Sp8, с закладкой зачатков плодолистика во время роста цветковых чешуй и черешков. (E–H) Замедление роста палео в osvil2 колосков. (E) Стадия Sp4, показывающая замедление зачатков палео (стрелка) по сравнению с ростом цветковых чешуй. (F) Стадия Sp6, с дегенеративным формированием палео и замедленным формированием зачатков тычинок на палео стороне. (G) Стадия Sp8, с замедленным ростом палео по сравнению с цветковой чешуей. (H) Стадия Sp6, демонстрирующая формирование зачатков тычинок (звездочка) в неправильном порядке. Дополнительные зачатки органов (острие стрелки) между палеей и тычинкой, увеличивающие размер цветочной мутовки. (И–Л) освил2 колосок с фенотипом махровых цветков. (I) Стадия Sp3, с более широкой меристемой цветка и двумя зачатками цветковых чешуй с обеих сторон. (J) Стадия Sp4, с двумя вырожденными зачатками палео, образующимися в середине двух цветковых чешуй. (K) Стадия Sp6, представляющая эктопические зачатки органов (острия стрелок), которые, вероятно, образуют дополнительные бледные пятна и лодикулы. (L) Стадия Sp6-7, с восемью зачатками тычинок. (M–P) osvil2 колосок с дополнительными колосковыми или чешуевидными органами. (M) Стадия Sp7, на которой число зачатков тычинок уменьшено до 5. Цветковые мутовки увеличены, а после дополнительной цветковой чешуи формируются вырожденные палео. (N) Стадия Sp4, с образованием дополнительных зачатков чешуи (стрелки). (O) Стадия Sp6, при которой образуются дополнительные леммы, а формирование внутренних органов задерживается. (P) Стадия Sp8, с дополнительной чешуей, образующейся после образования нормальных пустых чешуек и рудиментарных чешуек. Са, плодолистик; ДП, дегенеративное палео; EG, пустая чешуя; FM – цветочная меристема; L — лемма; Р, палеа; RG, рудиментарная чешуя. Шкала баров = 20 мкм.
У колосков osvil2 образование зачатков палео часто задерживалось на Sp4 (рис. 3Д). При Sp6 мутантные колоски демонстрировали вырожденные зачатки палео, часто вместе с задержкой развития внутренних органов цветка на палео стороне (рис. 3F, G).Эта задержка, по-видимому, вызвала снижение количества образующихся тычинок, а также аномальное развитие внутренних органов цветка (рис. 3H). ФМ были больше у некоторых osvil2 колосков, которые обычно сопровождали два зачатка цветковых чешуек и два вырожденных зачатка палео (рис. 3I,J). На Sp6 между цветковой чешуей и зачатками тычинок наблюдались дополнительные зачатки лодикул (рис. 3K). На этой стадии количество зачатков тычинок было изменено (рис. 3L, M), и иногда происходило повторное образование колосковых чешуй (рис. 3N).На дополнительных оборотах образовывались дополнительные колосковые чешуи и похожие на цветковые чешуи органы (рис. 3M–P). Эти данные показали, что OsVIL2 функционирует на ранних стадиях развития колосков.
Образец экспрессии
OsVIL2Паттерн экспрессии OsVIL2 анализировали с помощью ОТ-ПЦР. В вегетативных тканях ген был обнаружен в листьях проростков и в листовых пластинках взрослых растений (рис. 4А). Он также конститутивно экспрессировался в метелках на разных стадиях развития (рис. 4В).У зрелых колосков экспрессия была сильной в тычинках и плодолистиках и слабой в лодикулах и палеях (рис. 4С). Экспрессию также изучали на тканевом уровне с использованием промоторной области OsVIL2 , слитой с репортерным геном GUS (рис. 4D). Мы получили 13 растений, независимо трансформированных этой конструкцией промотора OsVIL2 — GUS . Эти линии демонстрировали схожий паттерн экспрессии, поэтому для дальнейшего анализа мы выбрали линию с самой высокой экспрессией GUS.Репортер GUS сильно экспрессировался в листьях, но не экспрессировался в корнях, как видно из анализов ОТ-ПЦР (фиг. 4E, F). Во время развития колосков репортер был обнаружен в базальных областях колосков (рис. 4G-J). В цветках Sp8 он был сильно выражен в пыльниках и в базальной области плодолистиков (рис. 4К–М). Этот предпочтительный для органов паттерн экспрессии согласовывался с паттерном, выявленным по данным qRT-PCR.
РИСУНОК 4. Характер экспрессии OsVIL2 в различных органах. (A–C) Анализ экспрессии ОТ-ПЦР. (A) Экспрессия в различных органах. (Б) Экспрессия в период раннего развития колосков. (C) Экспрессия в каждом цветочном органе внутри зрелых колосков. (D) Принципиальная схема ПОСВИЛ2 :: ГУС конструктив. (E–M) Наблюдение за линией захвата промотор-GUS OsVIL2. Экспрессия в листовой пластинке (E) , корень (F) , колосок Sp4 (G) , колосок Sp5 (H) , колосок Sp6 (I) , колосок Sp7 ( J) , колоски в 12-мм молодой метелке (K) , колоски в 50-мм метелке (L) , взрослый колосок (M) .EG, пустая чешуя; L — лемма; Р, палеа; RG, рудиментарная чешуя.
Транскриптомный анализ молодых метелок
Было проведеноанализа секвенирования РНК для идентификации генов, дифференциально экспрессирующихся в osvil2 во время раннего развития колосков. Были исследованы две стадии: 2-мм метелки, содержащие колоски в основном Sp4 или ранее, и 4-мм метелки, содержащие колоски в основном Sp7 или моложе. В образцах диаметром 2 мм 451 ген был активизирован (дополнительная таблица S2), а 606 генов были подавлены (дополнительная таблица S3) как минимум в два раза.В образцах диаметром 4 мм 548 генов были активизированы (дополнительная таблица S4), а 490 генов были подавлены (дополнительная таблица S5) как минимум в два раза. Перекрытие между 2- и 4-миллиметровыми образцами показало, что 330 генов были активизированы (дополнительная таблица S6), а 306 — подавлены (дополнительная таблица S7). В общей сложности 669 генов были активированы, а 790 генов были подавлены как минимум в два раза в обоих размерах (рис. 5).
РИСУНОК 5. Количество генов, дифференциально экспрессируемых в osvil2 молодых метелках. (A) Номер с повышающей регулировкой; (B) номер понижен.
Мы провели анализ ГО с использованием дифференциально экспрессируемых генов (дополнительная фигура S2). Наш GO-анализ показал, что Flower Development (GO:0009908) был значительно обогащен подавленными генами в 2-миллиметровых метелках, что означает, что регуляторные гены, участвующие в этом процессе, были подавлены в osvil2 на ранних стадиях. Термины для транскрипции (GO: 0006351) и регуляции транскрипции (GO: 0006355) также были сильно обогащены подавляющими генами обоих размеров метелки (дополнительные рисунки S2B, D).Среди факторов транскрипции многие гены семейства AP2 и MADS-box были значительно подавлены (дополнительная таблица S3). Поскольку некоторые гены в этих семействах играют важные функции в формировании SM и развитии органов цветка, их подавление в развитии метелок, вероятно, было причиной аномальных фенотипов колосков.
Транскриптомный анализ генов, участвующих в фазовом переходе меристемы или развитии органов колоска, показан в таблице 1. Гены, определяющие размер меристемы – FON1, FON4 и OsWUS (Suzaki et al., 2004; Чу и др., 2006; Мун и др., 2006; Nardmann, Werr, 2006) – почти одинаково экспрессировались у мутантов osvil2 . Среди генов, которые функционируют при переходе от меристемы соцветия (IM) к SM, транскриптомная частота APO1 была значительно снижена при обоих размерах метелки. APO1 , ортолог Arabidopsis UFO , подавляет более раннее превращение IM в SM и способствует пролиферации клеток в IM (Ikeda et al., 2005, 2007; Ikeda-Kawakatsu et al., 2009). Кроме того, APO1 регулирует идентичность цветочных органов и цветочную детерминированность путем усиления экспрессии C-функционального гена OsMADS3 (Ikeda et al., 2005). Однако мы обнаружили, что уровень экспрессии APO2 / RFL был повышен у наших мутантов osvil2 . Ген APO2 / RFL , ортолог Arabidopsis LFY , также подавляет переход от ИМ к СМ. Уменьшение его экспрессии уменьшает количество образующихся ветвей метелки и изменяет идентичность органов цветка (Rao et al., 2008; Икеда-Кавакацу и др., 2012).
ТАБЛИЦА 1. Уровни экспрессии генов, контролирующих развитие колосков.
Среди четырех генов RCN , гомологичных Arabidopsis TERMINAL FLOWER 1 (Nakagawa et al., 2002), экспрессия RCN4 была увеличена как минимум вдвое в мутантных колосках, в то время как экспрессия RCN1, RCN2 и RCN3 существенно не пострадали. Кроме того, нам не удалось обнаружить каких-либо существенных изменений в уровнях транскриптома для SNB и MFS , которые функционируют при переходе от SM к FM (таблица 1).Экспрессия была слегка повышена для FZP , гена, который ингибирует пазушную меристему в колоске и способствует FM путем усиления экспрессии B-функции ( OsMADS4 и OsMADS16 ), E-функции ( OsMADS1, OsMADS7 и OsMADS8 ) и AGL6-подобные ( OsMADS6 и OsMADS17 ) гены MADS-box (Komatsu et al., 2003; Bai et al., 2016).
Среди генов, необходимых для формирования пустых чешуй (Yoshida et al., 2009; Zhang et al., 2015), G1 и OsIG1 были значительно подавлены в молодых колосках по сравнению с osvil2 . Поскольку развитие пустых чешуй было аномальным и их количество было меньше у мутанта, за эти дефекты могла быть ответственна сниженная экспрессия генов. Более того, подавление REP1 , необходимое для развития palea (Yuan et al., 2009), может быть связано с дегенерацией palea у osvil2 .
Несколько генов MADS-box, которые функционируют в идентичности цветочных органов, были подавлены в колосках osvil2 .Экспрессия В-функции OsMADS4 и OsMADS16 была нарушена у мутанта, особенно в 2-мм колоске. С-функция OsMADS3, OsMADS58 и DL также подавлялась в мутантных колосках. Кроме того, экспрессия была значительно снижена для D-функции OsMADS13 и Е-функции OsMADS1, OsMADS6, OsMADS7 и OsMADS8 . Напротив, экспрессия генов A-функции OsMADS14, OsMADS15 и OsMADS18 существенно не изменилась, в то время как экспрессия OsMADS5 была увеличена.
Чтобы проверить наши данные секвенирования РНК, мы провели qRT-PCR, сосредоточив внимание на 13 генах MADS-box, которые контролируют идентичность и развитие цветочных органов (рис. 6). Среди них восемь ( OSMADS1, OSMADS3, OSMADS4, OSMADS6, OSMADS7, OSMADS13, OSMADS16 , и osmads16 и osmads58 ) были сокращены при выражении пяти ( osmads2, osmads5, osmads14, osmads15 и osmads34 ) существенно не изменился, согласно анализу транскриптома (рис. 6В).У мутантов osvil2 экспрессия всех восьми подавленных генов была значительно снижена на основании результатов qRT-PCR. Среди пяти, для которых анализ секвенирования РНК не показал значительных изменений, экспрессия была сходной между WT и osvil2 для трех, в то время как у двух других, OsMADS5 и OsMADS34 , была слегка повышена регуляция в мутантных метелках.
РИСУНОК 6. Проверка результатов секвенирования РНК с помощью qRT-PCR.Уровни транскрипта относятся к Ubi1 . (A) qRT-PCR анализ генов, участвующих в развитии колосков. (B) qRT-PCR анализ генов MADS-box, участвующих в развитии цветочных органов. (C) qRT-PCR анализ активированных генов, ранее идентифицированных в анализе секвенирования РНК.
Мы также выбрали восемь генов, экспрессия которых была подтверждена на основании нашего анализа транскриптома (рис. 6C). Эксперименты по проверке qRT-PCR показали, что их экспрессия была намного выше у мутантов osvil2 .Все эти результаты показали, что данные, полученные в результате анализа секвенирования РНК, надежны.
Обсуждение
В этом исследовании мы проанализировали аномальные фенотипы osvil2 и обнаружили, что они вариабельны во всех органах колосков. Кроме того, у мутанта была значительно изменена экспрессия важных регуляторных генов для развития колосков. Эти результаты продемонстрировали, насколько необходим OsVIL2 для формирования нормального паттерна органов во время формирования колосков, поскольку он модулирует правильную экспрессию тех генов, которые контролируют развитие и идентичность органов в колосках.
Колоски мутанта osvil2 продуцировали удлиненные пустые чешуйки, напоминающие цветковые чешуи. Гомеотическая трансформация пустых чешуй в цветковые чешуи описана также у мутантов, дефектных по G1, EG1, EG2, OsMADS34 и OsIG1 (Li et al., 2009; Yoshida et al., 2009; Gao et al., 2010). ; Cai et al., 2014; Zhang et al., 2015). Поскольку пустые чешуи не встречаются у колосков других злаков, происхождение и принадлежность пустых чешуй спорны (Yoshida, Nagato, 2011).Удлиненный фенотип пустых чешуй osvil2 подтверждает идею о том, что пустые чешуи представляют собой вырожденные чешуи стерильных цветков (Arber, 1934). Экспрессия генов пустых колосковых чешуек – G1 и OsIG1 – была значительно ниже в развивающихся метелках osvil2 , чем у WT, что позволяет предположить, что OsVIL2 функционирует выше G1 и Os4IG1 до указать стерильную идентичность леммы.
Другим фенотипом osvil2 колосков было образование вырожденных палеа.Этот дефект возник в основном в палеотеле, а не в краевой области. Кроме того, средняя часть зачатков палео часто не росла, что приводило к расщеплению палео. Эти фенотипы аналогичны фенотипам, зарегистрированным для мутантов, дефектных в REP1, OsMADS15, MFS1, DP1 или OsIG1 (Yuan et al., 2009; Wang et al., 2010; Jin et al., 2011; Ren et al. ., 2013; Чжан и др., 2015). Экспрессия REP1, DP1 и OsIG1 была снижена у мутантов osvil2 , что позволяет предположить, что эти гены связаны с дефектами palea.У арабидопсиса гены VIL функционируют во время яровизации и цветения (Sung, Amasino, 2004; Sung et al., 2006; Greb et al., 2007). Наше наблюдение, что рис VIL участвует в развитии колосков, указывает на диверсифицированную функцию семейства генов. Поскольку колосок представляет собой уникальную структуру соцветия травы, будет интересно изучить, сохраняется ли функция OsVIL2 у других видов трав.
Наиболее значимым фенотипом osvil2 было увеличение количества всех органов цветка.Этот фенотип был подобен таковому у мутантов, дефектных в FON4 , ортологе Arabidopsis CLV3 (Chu et al., 2006). При этом количество цветочных органов во внутренних мутовках больше зависит от колосков. У мутантов fon4 количество плодолистиков может увеличиваться до 10, тогда как у osvil2 это число возрастает до двух. Количество тычинок также увеличивается до 10 у fon4 по сравнению с 8 у osvil2 . Гомеотическая конверсия пустых чешуй в цветковые чешуи характерна как для мутантов fon4 , так и osvil2 .Мы отметили, что экспрессия FON4 была снижена в развивающихся метелках osvil2 , предполагая, что этот ген функционирует ниже по течению от OsVIL2 .
Идентичность цветочных органов регулируется многочисленными генами, включая некоторые гены MADS-box (Zhang and Yuan, 2014). Мы наблюдали, что несколько генов MADS-box по-разному экспрессировались у osvil2 . Изменение экспрессии этих цветочных гомеозисных генов могло быть ответственно за аномальное развитие цветочных органов у мутанта.
Это исследование показало, что OsVIL2 влияет на различные аспекты развития колосков, контролируя различные гены, важные для развития колосков. Поскольку OsVIL2 функционирует вместе с PRC2, который подавляет хроматин-мишень, мы ожидали обнаружить, что экспрессия прямых мишеней будет выше у мутантов osvil2 . Вместо этого уровни транскрипции для большинства регуляторных генов, которые контролируют количество или идентичность органов, были снижены, в то время как экспрессия была немного увеличена для RCN4, OsMADS5 и OsMADS34 .Это означает, что они могут быть прямыми мишенями OsVIL2-PRC2. Хотя функция RCN4 остается неизвестной, сверхэкспрессия RCN1 и RCN2 может приводить к сильно разветвленным метелкам, указывая на то, что они также играют роль в подавлении судьбы цветка (Nakagawa et al., 2002). В более раннем функциональном исследовании OsEMF2b было предсказано, что OsMADS34 является прямым геном-мишенью Os EMF2b (Conrad et al., 2014). Поскольку OsEMF2b взаимодействует с OsVIL2 (Yang et al., 2013), мутации OsVIL2 также могут влиять на гены MADS-box.
Вклад авторов
GA организовал все это исследование. GA, HY, JY, WL и DZ разработали исследование. HY, JY, WL и DZ провели эксперименты и проанализировали данные. HY и GA написали рукопись. Все авторы прочитали и одобрили рукопись.
Финансирование
Эта работа была поддержана грантом Совместной исследовательской программы по развитию сельскохозяйственной науки и техники, Управление по развитию сельских районов, Южная Корея (проект №.PJ013210) для GA.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят Кёнсука Ана за обработку запаса семян и Присциллу Лихт за редактирование содержания статьи на английском языке.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2018.00102/full#supplementary-material
РИСУНОК S1 | Фенотипы освил2-2 колоски. (А) Аномальное рудиментарное образование чешуи у освил2-2 колоска. (Б) освил2-2 колосок с удлиненными пустыми чешуями. (C) osvil2-2 колосок с дополнительными чешуевидными органами, обозначенными стрелками. (Г) освиль2-2 колосок с вырожденной палеей. (E) Фенотип двойных цветков. (F) Дополнительные органы цветка. (G) Дополнительная лодикула и образование удлиненной лодыжки. (H) Аномальный плодолистик с увеличенным числом рылец и недифференцированной клеточной массой. (I) Лодикуло-тычиночный мозаичный орган в osvil2-2 . Са, плодолистик; ЭЭГ — удлиненная пустая чешуя; Эло, удлиненный лодикула; L — лемма; Лодикула; Р, палеа; мкм, недифференцированная клеточная масса. Шкала баров = 1 мм.
РИСУНОК S2 | Генеонтологический анализ дифференциально экспрессируемых генов osvil2 . (A) Обогащение активированных генов в 2-мм метелке. (B) Обогащение подавленных генов в 2-мм метелке. (C) Обогащение активированных генов в 4-мм метелке. (D) Обогащение подавленных генов в 4-мм метелке.
ТАБЛИЦА S1 | Последовательности праймеров, использованных в этом исследовании.
ТАБЛИЦА S2 | Гены активировались в 2-миллиметровых молодых метелках osvil2 .
ТАБЛИЦА S3 | Гены подавлены в 2-миллиметровых молодых метелках osvil2 .
ТАБЛИЦА S4 | Гены активировались в 4-миллиметровых молодых метелках osvil2 .
ТАБЛИЦА S5 | Гены подавлены в 4-мм молодых метелках osvil2 .
ТАБЛИЦА S6 | Гены активировались как в 2-, так и в 4-мм молодых метелках osvil2 .
ТАБЛИЦА S7 | Гены подавлены как в 2-, так и в 4-мм молодых метелках osvil2 .
Сноски
- http://www.ncbi.nlm.nih.gov/
- http://www.uniprot.org/
- http://www.geneontology.org/
Каталожные номера
Ан Г., Эберт П. Р., Митра А. и Ха С. Б. (1989). «Бинарные векторы», в The Plant Molecular Biology Manual , eds SB Gelvin and R.A. Schilperoort (Dordrecht: Kluwer Academic Publisher), 1–19.
Академия Google
Арбер, А. (1934). Злаки: исследование злаков, бамбука и трав .Кембридж: Издательство Кембриджского университета.
Академия Google
Бай, X., Хуан, Ю., Мао, Д., Вэнь, М., Чжан, Л., и Син, Ю. (2016). Регуляторная роль ФЗП в детерминации ветвления метелок и образования колосков у риса. науч. Респ. 6:19022. дои: 10.1038/srep19022
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боммерт П., Сато-Нагасава Н., Джексон Д. и Хирано Х.Ю. (2005). Генетика и эволюция развития соцветий и цветков у злаков. Физиол клеток растений. 46, 69–78. doi: 10.1093/pcp/pci504
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cai, Q., Yuan, Z., Chen, M., Yin, C., Luo, Z., Zhao, X., et al. (2014). Жасмоновая кислота регулирует развитие колосков у риса. Нац. коммун. 5:3476. дои: 10.1038/ncomms4476
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Цао, Р., Ван, Л., Ван, Х., Ся, Л., Эрджумент-Бромейдж, Х., Темпст, П., и соавт.(2002). Роль метилирования лизина 27 гистона h4 в молчании группы Polycomb. Наука 298, 1039–1043. doi: 10.1126/наука.1076997
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Chu, H., Qian, Q., Liang, W., Yin, C., Tan, H., Yao, X., et al. (2006). Ген FLORAL ORGAN NUMBER4 , кодирующий предполагаемый ортолог Arabidopsis CLAVATA3, регулирует размер апикальной меристемы риса. Завод физиол. 142, 1039–1052. дои: 10.1104/стр.106.086736
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Конрад, Л.Дж., Хандай, И., Джонсон, К., Гидердони, Э., Ан, Г., Виджайрагхаван, У., и др. (2014). Ген группы поликомб EMF2B необходим для поддержания детерминированности цветочной меристемы у риса. Завод J. 80, 883–894. doi: 10.1111/tpj.12688
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чермин Б., Мелфи Р., Маккейб Д., Зейтц В., Имхоф, А., и Пирротта, В. (2002). Энхансер комплексов Zeste/ESC дрозофилы обладает активностью метилтрансферазы гистона h4, которая маркирует хромосомные сайты Polycomb. сотовый 111, 185–196. doi: 10.1016/S0092-8674(02)00975-3
Полнотекстовая перекрестная ссылка | Академия Google
Ган, Э. С., Хуанг, Дж., Ито, Т. (2013). Функциональная роль модификации гистонов, ремоделирования хроматина и микроРНК в развитии цветков Arabidopsis . Междунар. Рев. ячейка Мол.биол. 305, 115–161. дои: 10.1016/B978-0-12-407695-2.00003-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гао X., Лян В., Инь К., Цзи С., Ван Х., Су Х. и др. (2010). Ген SEPALLATA -подобный OsMADS34 необходим для развития соцветий и колосков риса. Завод физиол. 153, 728–740. doi: 10.1104/стр.110.156711
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гудрич, Дж., Puangsomlee, P., Martin, M., Long, D., Meyerowitz, E.M., and Coupland, G. (1997). Ген группы Polycomb регулирует экспрессию гомеозисного гена у Arabidopsis . Природа 386, 44–51. дои: 10.1038/386044a0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Greb, T., Mylne, J.S., Crevillen, P., Geraldo, N., An, H., Gendall, A.R., et al. (2007). Белок пальца PHD VRN5 функционирует в эпигенетическом сайленсинге Arabidopsis FLC . Курс. биол. 17, 73–78. doi: 10.1016/j.cub.2006.11.052
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хенниг Л., Таранто П., Вальзер М., Шенрок Н. и Груиссем В. (2003). Arabidopsis MSI1 необходим для эпигенетического поддержания репродуктивного развития. Разработка 130, 2555–2565. doi: 10.1242/dev.00470
Полнотекстовая перекрестная ссылка | Академия Google
Икеда К., Ито М., Нагасава Н., Кёзука Дж.и Нагато, Ю. (2007). Rice ABERRANT PANICLE ORGANIZATION 1 , кодирующий белок F-box, регулирует судьбу меристемы. Завод Ж. 51, 10:30–10:40. doi: 10.1111/j.1365-313X.2007.03200.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Икеда, К., Нагасава, Н., и Нагато, Ю. (2005). АБЕРРАНТНАЯ ОРГАНИЗАЦИЯ МЕТЕЛОК 1 временно регулирует идентичность меристем у риса. Дев. биол. 282, 349–360. doi: 10.1016/j.ydbio.2005.03.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Икеда, К., Сунохара Х. и Нагато Ю. (2004). Ход развития соцветия и колоска риса. Порода. науч. 54, 147–156. doi: 10.1270/jsbbs.54.147
Полнотекстовая перекрестная ссылка | Академия Google
Икеда-Кавакацу, К., Маекава, М., Идзава, Т., Ито, Дж. И., и Нагато, Ю. (2012). ABERRANT PANICLE ORGANIZATION 2/RFL , рисовый ортолог Arabidopsis LEAFY , подавляет переход от меристемы соцветия к меристеме цветка посредством взаимодействия с APO1 . Завод Ж. 69, 168–180. doi: 10.1111/j.1365-313X.2011.04781.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Икэда-Кавакацу, К., Ясуно, Н., Ойкава, Т., Иида, С., Нагато, Ю., Маекава, М., и другие. (2009). Уровень экспрессии ABERRANT PANICLE ORGANIZATION1 определяет форму соцветия риса посредством контроля пролиферации клеток в меристеме. Завод физиол. 150, 736–747. doi: 10.1104/стр.109.136739
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чон, Дж.S., Chung, Y.Y., Lee, S., Yi, G.H., Oh, B.G. и An, G. (1999). Выделение и характеристика специфичного для пыльников гена RA8 из риса ( Oryza sativa L.). Завод Мол. биол. 39, 35–44. дои: 10.1023/A:1006157603096
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чон, Дж. С., Джанг, С., Ли, С., Нам, Дж., Ким, К., Ли, С. Х., и др. (2000а). листовая оболочка sterile1 представляет собой гомеотическую мутацию в гене коробки риса MADS, влияющую на развитие цветков риса. Растительная клетка 12, 871–884. doi: 10.1105/tpc.12.6.871
Полнотекстовая перекрестная ссылка | Академия Google
Чон, Дж. С., Ли, С., Юнг, К. Х., Джун, С. Х., Чон, Д. Х., Ли, Дж., и др. (2000б). Инсерционный мутагенез Т-ДНК для функциональной геномики риса. Завод J. 22, 561–570. doi: 10.1046/j.1365-313x.2000.00767.x
Полнотекстовая перекрестная ссылка | Академия Google
Чон, Д. Х., Ан, С., Канг, Х. Г., Мун, С., Хан, Дж. Дж., Парк, С., и др. (2002).Инсерционный мутагенез Т-ДНК для мечения активации в рисе. Завод физиол. 130, 1636–1644. doi: 10.1104/стр.014357
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Jin, Y., Luo, Q., Tong, H., Wang, A., Cheng, Z., Tang, J., et al. (2011). Ген AT-hook необходим для формирования палеа и контроля количества цветочных органов у риса. Дев. биол. 359, 277–288. doi: 10.1016/j.dbio.2011.08.023
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кац, А., Олива М., Москуна А., Хаким О. и Охад Н. (2004). Поликомб-белки FIE и CURLY LEAF взаимодействуют в регуляции экспрессии гена гомеобокса во время развития спорофита. Завод J. 37, 707–719. doi: 10.1111/j.1365-313X.2003.01996.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Комацу М., Чуджо А., Нагато Ю., Симамото К. и Кёзука Дж. (2003). FRIZZY PANICLE необходим для предотвращения образования пазушных меристем и установления идентичности цветочных меристем в колосках риса. Разработка 130, 3841–3850. doi: 10.1242/dev.00564
Полнотекстовая перекрестная ссылка | Академия Google
Ли, Д.Ю., и Ан, Г. (2012). Два гена семейства AP2, SUPERNUMERARY BRACT ( SNB ) и OsINDETERMINATE SPIKELET 1 ( OsIDS1 ), синергетически контролируют архитектуру соцветия и формирование цветочной меристемы у риса. Завод J. 69, 445–461. doi: 10.1111/j.1365-313X.2011.04804.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли, Д.Ю., Ли Дж., Мун С., Парк С.Ю. и Ан Г. (2007). Ген гетерохронности риса SUPERNUMERARY BRACT регулирует переход от меристемы колоска к меристеме цветка. Завод Ж. 49, 64–78. doi: 10.1111/j.1365-313X.2006.02941.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ленхард М., Бонерт А., Юргенс Г. и Лаукс Т. (2001). Прекращение поддержания стволовых клеток в цветочных меристемах Arabidopsis за счет взаимодействия между WUSCHEL и AGAMOUS . Сотовый 105, 805–814. doi: 10.1016/S0092-8674(01)00390-7
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Li, H., Liang, W., Jia, R., Yin, C., Zong, J., Kong, H., et al. (2010). AGL6 -подобный ген OsMADS6 регулирует идентичность цветочных органов и меристем у риса. Сотовые Res. 20, 299–313. doi: 10.1038/cr.2009.143
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли Х., Сюэ Д., Гао З., Yan, M., Xu, W., Xing, Z., et al. (2009). Предполагаемый ген липазы EXTRA GLUME1 регулирует как судьбу пустых чешуй, так и развитие колосков у риса. Завод J. 57, 593–605. doi: 10.1111/j.1365-313X.2008.03710.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Луо, М., Платтен, Д., Чаудхури, А., Пикок, В.Дж., и Деннис, Э.С. (2009). Экспрессия, импринтинг и эволюция гомологов риса генов группы поликомб. Мол. Завод 2, 711–723.doi: 10.1093/mp/ssp036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mayer, K.F., Schoof, H., Haecker, A., Lenhard, M., Jürgens, G., and Laux, T. (1998). Роль WUSCHEL в регуляции судьбы стволовых клеток в меристеме побегов Arabidopsis . сотовый 95, 805–815. doi: 10.1016/S0092-8674(00)81703-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мун С., Юнг К. Х., Ли Д. Э., Ли Д. Ю., Ли Дж., Ан, К., и др. (2006). Ген FON1 риса контролирует вегетативное и репродуктивное развитие, регулируя размер апикальной меристемы побега. Мол. Ячейки 21, 147–152.
Реферат PubMed | Академия Google
Moon, Y.H., Chen, L., Pan, R.L., Chang, H.S., Zhu, T., Maffeo, D.M., et al. (2003). генов EMF поддерживают вегетативное развитие, подавляя программу цветения у Arabidopsis . Растительная клетка 15, 681–693. дои: 10.1105/тпк.007831
Полнотекстовая перекрестная ссылка | Академия Google
Мозгова И., Келер К. и Хенниг Л. (2015). Держите ворота закрытыми: функции поликомбного репрессивного комплекса PRC2 в развитии. Завод Ж. 83, 121–132. doi: 10.1111/tpj.12828
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Müller, J., Hart, C.M., Francis, N.J., Vargas, M.L., Sengupta, A., Wild, B., et al. (2002). Гистонметилтрансферазная активность комплекса репрессора группы Drosophila Polycomb. сотовый 111, 197–208. doi: 10.1016/S0092-8674(02)00976-5
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Накагава, М., Симамото, К., и Киозука, Дж. (2002). Сверхэкспрессия RCN1 и RCN2 , гомологов риса TERMINAL FLOWER 1/CENTRORADIALIS , вызывает задержку фазового перехода и изменение морфологии метелки у риса. Завод J. 29, 743–750. doi: 10.1046/j.1365-313X.2002.01255.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нардманн, Дж.и Верр, В. (2006). Ниша стволовых клеток побега у покрытосеменных: паттерны экспрессии ортологов WUS у риса и кукурузы предполагают серьезные модификации в ходе эволюции одно- и двудольных растений. Мол. биол. Эвол. 23, 2492–2504. doi: 10.1093/molbev/msl125
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Омори С., Кимидзу М., Сугита М., Мияо А., Хирочика Х., Учида Э. и др. (2009). MOSAIC FLORAL ORGANS1 , AGL6 -подобный ген коробки MADS, регулирует идентичность цветочных органов и судьбу меристемы риса. Растительная клетка 21, 3008–3025. doi: 10.1105/tpc.109.068742
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Прасада, К., Парамешваран, С., и Виджайрагхаван, У. (2005). OsMADS1 , фактор MADS-box риса, контролирует дифференцировку определенных типов клеток в лепестках и палеях и является регулятором раннего действия внутренних цветочных органов. Завод Ж. 43, 915–928. doi: 10.1111/j.1365-313X.2005.02504.x
Реферат PubMed | Полнотекстовая перекрестная ссылка
Рао, Н.Н., Прасад К., Кумар П.Р. и Виджайрагхаван У. (2008). Особая регуляторная роль RFL , гомолога риса LFY , в определении времени цветения и строения растения. Проц. Натл. акад. науч. США 105, 3646–3651. doi: 10.1073/pnas.0700
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рен Д., Ли Ю., Чжао Ф., Санг Х., Ши Дж., Ван Н. и др. (2013). MULTI-FLORET SPIKELET1 , который кодирует белок AP2/ERF, определяет судьбу меристемы колоска и идентичность стерильной цветочной чешуи у риса. Завод физиол. 162, 872–884. doi: 10.1104/PP.113.216044
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Санг, X., Ли, Ю., Луо, З., Рен, Д., Фанг, Л., Ван, Н., и др. (2012). CHIMERIC FLORAL ORGANS1 , кодирующий специфичный для однодольных растений белок MADS box, регулирует идентичность цветочных органов у риса. Завод физиол. 160, 788–807. doi: 10.1104/pp.112.200980
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вс, Б., Looi, L.S., Guo, S., He, Z., Gan, E.S., Huang, J., et al. (2014). Механизм синхронизации, зависящий от клеточного деления, запускается выселением Polycomb в стволовых клетках растений. Наука 343:1248559. doi: 10.1126/наука.1248559
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сунь, Б., Сюй, Ю., Нг, К. Х., и Ито, Т. (2009). Временной механизм поддержания и дифференцировки стволовых клеток в меристеме цветков Arabidopsis . Гены Дев. 23, 1791–1804 гг.doi: 10.1101/gad.1800409
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сунг, С., Шмитц, Р. Дж., и Амасино, Р. М. (2006). Белок пальца PHD, участвующий как в путях яровизации, так и в путях фотопериода у Arabidopsis . Гены Дев. 20, 3244–3248. doi: 10.1101/gad.1493306
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Судзаки Т., Сато М., Асикари М., Миёси М., Нагато Ю. и Хирано Х.Ю.(2004). Ген НОМЕР ЦВЕТОЧНОГО ОРГАНА 1 регулирует размер цветочной меристемы риса и кодирует киназу богатого лейцином повторного рецептора, ортологичную Arabidopsis CLAVATA1. Разработка 131, 5649–5657. doi: 10.1242/dev.01441
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ван, К., Тан, Д., Хонг, Л., Сюй, В., Хуанг, Дж., Ли, М., и другие. (2010). DEP и AFO регулируют репродуктивную функцию риса. ПЛОС Жен. 6:e1000818. doi: 10.1371/journal.pgen.1000818
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вей, Дж., Чой, Х., Джин, П., Ву, Ю., Юн, Дж., Ли, Ю.С., и др. (2016). Ген гомеобокса типа GL2 Roc4 у риса способствует продолжительности цветения преимущественно при длинном дне за счет репрессии Ghd7 . Растениевод. 252, 133–143. doi: 10.1016/j.plantsci.2016.7.012
Полнотекстовая перекрестная ссылка | Академия Google
Се, С., Чен М., Пей Р., Оуян Ю. и Яо Дж. (2015). OsEMF2b действует как регулятор перехода цветения и идентичности цветочных органов, опосредуя отложение h4K27me3 в OsLFL1 и OsMADS4 у риса. Завод Мол. биол. 33, 121–132. doi: 10.1007/s11105-014-0733-1
Полнотекстовая перекрестная ссылка | Академия Google
Ян, Дж., Ли, С., Ханг, Р., Ким, С.Р., Ли, Ю.С., Цао, X., и др. (2013). OsVIL2 функционирует с PRC2, вызывая цветение путем подавления OsLFL1 у риса. Завод J. 73, 566–578. doi: 10.1111/tpj.12057
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Юн, Дж., Чо, Л. Х., Ким, С. Л., Чой, Х., Кох, Х. Дж., и Ан, Г. (2014). Ген гомеобокса типа BEL1 SH5 индуцирует осыпание семян за счет усиления развития зоны абсциссии и ингибирования биосинтеза лигнина. Завод J. 79, 717–728. doi: 10.1111/tpj.12581
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Йошида, А., Судзаки Т., Танака В. и Хирано Х.Ю. (2009). Гомеозисный ген длинная стерильная цветковая чешуя ( G1 ) определяет идентичность стерильной цветочной чешуи в колоске риса. Проц. Натл. акад. науч. США 106, 20103–20108 гг. doi: 10.1073/pnas.0
6106Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Йошида Н., Янаи Ю., Чен Л., Като Ю., Хирацука Дж., Мива Т. и др. (2001). EMBRYONIC FLOWER2, новый гомолог белка группы поликомб, опосредует развитие побегов и цветение у Arabidopsis . Растительная клетка 13, 2471–2481. doi: 10.1105/tpc.010227
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Юань З., Гао С., Сюэ Д. В., Луо Д., Ли Л. Т., Дин С. Ю. и др. (2009). RETARDED PALEA1 контролирует развитие палеа и зигоморфию цветков риса. Завод физиол. 149, 235–244. doi: 10.1104/стр.108.128231
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжан Д. и Юань З. (2014).Молекулярный контроль развития стекловидного соцветия. год. Преподобный Завод Физиол. 65, 553–578. doi: 10.1146/annurev-arplant-050213-040104
Реферат PubMed | Полнотекстовая перекрестная ссылка
Zhang, J., Tang, W., Huang, Y., Niu, X., Zhao, Y., Han, Y., et al. (2015). Понижающая регуляция LBD -подобного гена, OsIG1 , приводит к возникновению необычных двойных семязачатков и аномалиям развития различных цветочных органов и мегагаметофитов у риса. Дж.Эксп. Бот. 66, 99–112. дои: 10.1093/jxb/eru396
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжао, С. К., Ху, Дж., Го, Л. Б., Цянь, К., и Сюэ, Х. В. (2010). Наклон листа риса2, VIN3-подобный белок, регулирует угол листа посредством модулирования клеточного деления воротничка. Сотовые Res. 20, 935–947. doi: 10.1038/cr.2010.109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Растения | Колосок, другой «цветок» в рисе
Описательный абзац.Растительная клетка https://doi.org/10.1105/tpc.18.00682
Хуэй Чжуан и Юньфэн Ли
Предыстория: Колосок, который часто ошибочно принимают за «цветок», на самом деле является соцветием и может образовывать один или несколько соцветий у некоторых видов трав. Это означает, что можно было бы увеличить количество соцветий (зерен) в колоске и повысить урожайность у видов, которые в настоящее время дают только одноцветковый колосок. У риса колосок содержит один фертильный верхушечный цветочек, одну пару стерильных цветковых чешуй и одну пару рудиментарных колосковых чешуй.Хотя рис дикого типа дает только один цветочек в колоске, потеря детерминированности меристем колосков или восстановление двух «боковых цветков», которые теперь считаются предшественниками «стерильных цветковых чешуй», может способствовать увеличению количества цветков. соцветий в колоске. Однако для этого необходимо более глубокое понимание генетического и молекулярного механизма развития колосков.
Вопросы: Какой фактор является ключевым в развитии колосков?
Находки: Мы идентифицировали новый репрессор транскрипции, названный NSG1 , который кодирует белок цинкового пальца C2h3 и играет ключевую роль в регуляции идентичности боковых органов колоска.В мутантном колоске nsg1-1 LHS1 , DL и MFO1 эктопически экспрессировались в двух или более органах, включая рудиментарную чешуйку, стерильную цветковую чешую, палео, лодикулу и тычинку, тогда как G7 был подавляется в рудиментарной чешуе и стерильной чешуе. Кроме того, белок NSG1 был способен связываться с регуляторными областями LHS1 , а затем рекрутировать корепрессорные TPR для подавления экспрессии путем снижения уровней ацетилирования гистонов в хроматине.Наше исследование показывает, что большинство генов, контролирующих развитие колосков, регулируются вышестоящими промоторными областями и факторами транскрипции.
Следующие шаги: Мы проведем более глубокий анализ, чтобы идентифицировать гены развития колосков и их молекулярную регуляторную сеть у риса. Кроме того, мы проведем молекулярно-дизайнерскую селекцию «трехцветковых колосков» с использованием нескольких генов развития колосков, в том числе NSG1 и LF1 , которые были идентифицированы нашей группой в 2017 г. и индуцировали образование более поздних цветков на месте стерильных. лемма (Чжан и др., 2017 PNAS).
Hui Zhuang, Hong-Lei Wang, Ting Zhang, Xiao-Qin Zeng, Huan Chen, Zhong-Wei Wang, Jun Zhang, Hao Zheng, Jun Tang, Ying-Hua Ling, Zheng-Lin Yang, Guang-Hua He and Yun-Feng Li. (2019). NONSTOP GLUMES 1 Encodes a C2h3 Zinc Finger Protein that Regulates Spikelet Development in Rice. Plant Cell. https://doi.org/10.1105/tpc.18.00682.
Key words: TFs, C2h3 zinc finger, spikelet development, rice
(PDF) Understanding spikelet orientation in Paniceae (Poaceae)
13
May 2010] Zanotti et al.— Ориентация колосков у Paniceae (Poaceae)
Giraldo-Cañas, D. 2007 . An álisis filogené tico del gé nero neotropi-
cal Axonopus (Poaceae: Panicoideae: Paniceae) con base en carac-
teres morphol — gicos y anat — micos. Revista Institucional Universidad
Tecnol ó gica del Choc ó D.L.C. 26: 9 – 27.
Джуссани, Л. М., Дж. Х. Кота-Санчес, Ф. О. Сулоага и Э.А.
Келлог. 2001 . Молекулярная филогения подсемейства трав
Panicoideae (Poaceae) показывает множественное происхождение фотосинтеза C
4 .
Американский журнал ботаники 88: 1993–2001.
Го Мез-Мартинес, Р. и А. Калхэм. 2000 . Филогения подсемейства
ily Panicoideae с акцентом на трибу Paniceae: данные
области хпДНК trnL-F.В SWL Jacobs and J. Everett [eds.],
Grasses: Систематика и эволюция, 136–140. CSIRO Publishing,
Коллингвуд, Виктория, Австралия.
GPWG [Рабочая группа по филогении трав] . 2001 . Филогения и
подсемейная классификация злаков (Poaceae). Летопись ботанического сада
Миссури 88: 373–457.
Хсу, CC 1965. Классификация Panicum (Gramineae) и его ал-
верна, с особой ссылкой на признаки лодикулы, стилевой основы и
леммы.Журнал факультета естественных наук Токийского университета, раздел
3: Ботаника 9: 43–150.
Келлог, Э. А. 2000. Молекулярная и морфологическая эволюция
Andropogoneae. В SWL Jacobs and J. Everett [eds.], Grasses:
Систематика и эволюция, 149–158. CSIRO Publishing, Collingwood,
Victoria, Australia.
Келлог, Э. А. 2006. Помимо таксономии: перспективы понимания морфологического разнообразия злаков (Poaceae).Дарвиниана 44: 7-17.
Kellogg, E.A., K.M. Hiser, and A.N. Doust. 2004 . Таксономия, филогения и развитие соцветий рода Ixophorus (Panicoideae:
Poaceae). Международный журнал наук о растениях 165: 1089–1105.
Ле Ру, Л. Г. и Э. А. Келлогг. 1999 . Развитие цветка и
образование однополых колосков у Andropogoneae (Poaceae).
Американский журнал ботаники 86: 354 – 366.
Лю, К., П. М. Петерсон, Дж. Т. Колумб, Н. Чжао, Г. Хао и Д.
Чжан. 2007 . Диверсификация соцветий в «кладе пальчатого проса»
(Chloridoideae, Poaceae): сравнение молекулярной филогении и де-
морфологии покровов. Американский журнал ботаники 94: 1230–1247.
Морроне, О., Ф. О. Сулоага и Э. Карбон. 1995 . Revisión del
grupo Racemosa del gé nero Paspalum (Poaceae: Panicoideae:
Paniceae). Анналы Ботанического сада Миссури 82: 82–116.
Орр, А. Р., К. Маллен, Д. Клаахсен и М. Д. Сандберг. 2002 .
Развитие соцветия высокогорного однолетнего мексиканского теосинте
(Poaceae).Американский журнал ботаники 89: 1730-1740.
Орр, А. Р. и М. Д. Сандберг. 2004 . Развитие соцветий у
нового теосинте: Zea nicaraguensis (Poaceae). Американский журнал
Ботаника 91: 165–173.
Reinheimer, R., R. Pozner, and A.C. Vegetti. 2005 . Соцветие,
колосковое и цветочное развитие у Panicum maximum и Urochloa
plantaginea (Poaceae).Американский журнал ботаники 92: 565–575.
Райнхеймер Р., Ф. О. Сулоага, А. К. Вегетти и Р. Познер. 2009 .
Диверсификация развития соцветий в кладе PCK
(Poaceae: Panicoideae: Paniceae). Американский журнал ботаники 96:
549 – 564.
Руа, Г. Х. 1993. Синфлоресценция Paspalidium rarum (Poaceae)
и альтернативная гипотеза эволюции соцветий некоторых злаковых
.Австралийский журнал ботаники 6: 261–267.
Руа, Г. Х. и Ф. Веберлинг. 1995 . Форма роста и структура соцветия
Paspalum L. (Poaceae, Paniceae): сравнительно-
морфологический подход. Beiträge zur Biologie Pflanzen 69: 363–431.
Рузин С.Е. 1999 . Растительная микротехника и микроскопия. Oxford
University Press, Нью-Йорк, Нью-Йорк, США.
Сендульский Т. и Бурман А.Г. 1978 . Виды Paspalum из
Серра-ду-Сип ó (I): Вклад в изучение бразильских Poaceae.
Revista Brasileira Bot nica 1 : 1 – 15 .
Соуза-Чис, Т. Т., Л. Эсси, Г. Х. Руа, Дж. Ф. М. Валлс и Р. Б.
Миз. 2006 . Предварительный подход к филогении рода
Paspalum (Poaceae).Генетика 126: 15–32.
Stür, WW 1986. Репродуктивное развитие верхушки Brachiaria
decumbens Stapf. Анналы ботаники 58: 569-575.
Томлинсон, П. Б. 1971. Вершина побега и его дихотомическое разветвление
у пальмы Нипа. Анналы ботаники 35: 865–879.
Томлинсон, П. Б. и У. Послушны. 1977 . Особенности дихотомии
вершин у Flagellaria indica (однодольные).Американский журнал
Ботаника 64: 1057-1065.
Вегетти, AC 1987. An á lisis tipol — gico de la inflorescencia en Paspalum
(Poaceae). Курциана 19: 155–160.
Vollbrecht, E., P.S. Springer, L. Goh, E.S. Buckler, and R.
Martienssen. 2005 . Архитектура систем цветочных ветвей кукурузы
и родственных трав.Природа 436: 1119 – 1126.
Вебстер, Р. Д. 1988. Роды североамериканских Paniceae (Poaceae:
Panicoideae). Систематическая ботаника 13: 576–609.
Вебстер, Р. Д. 1992. Значение признаков и родовое сходство у
Paniceae (Poaceae: Panicoideae) . Сида 15: 185-213.
Ву, Х. и П. Макстин. 2007 . Роль транспорта ауксина во время развития цветения кукурузы (Zea mays, Poaceae).Американский
Журнал ботаники 94: 1745-1755.
Ву, X., А. Скирпан и П. Макстин. 2009 . Супрессор сидячих
колосков1 функционирует в рамозном пути, контролирующем детерминацию меристемы
кукурузы. Физиология растений 149: 205-219.
Сулоага, Ф. О. и О. Морроне. 2005 . Revisi ó n de las especies de
Paspalum para Amé rica del Sur Austral (Аргентина, Боливия, Sur
del Brasil, Чили, Парагвай и Уругвай).Монографии в Систематике
Ботаника из Ботанического сада Миссури 102: 1 – 297.
Сулоага, Ф. О., О. Морроне и Л. М. Джуссани. 2000 . Кладистический
анализ Paniceae: предварительный подход. В SWL Jacobs
и J. Everett [eds.], Grasses: Systematics and evolution, 123–135.
CSIRO Publishing, Коллингвуд, Виктория, Австралия.
Paspalum simplex Morong ex Britton — Аргентина: Guaglianone, E.Р. и др.
281 ; Льюис, PJ 639; Морроне О. и Джуссани Л. М. 5794*; Педерсен,
Т. М. 8283 ; Quarín, C. 2955; Ротман, А.Д. и другие. 245 ; Schinini, A. и
Martí nez Crovetto, R . 12310; Шульц, А. Г. 1172 9. Боливия: Киллин,
Т. 1722. Парагвай: Буркарт, А. 18302; Буркарт, А. 18404; Мерелес, Ф.
и Рамелла, Л. 2611; Сулоага, Ф. О. и Морроне, О. 7328. Уругвай:
Розенгуртт, Б.Б-7142.
Paspalum stellatum Humb. & Бонпл. ex Flü gg é — Аргентина: Cabrera, AL
28254; Краповицкас, А. и др. . 25202 ; Морроне О. и Джуссани Л. М.
5803*; Zuloaga, F. O. et al. 910 . Боливия: Киллин, Т. 1952. Бразилия: Irwin,
HS и Soderstrom, TR 5163, Pedersen, TM 15713. Парагвай:
Burkart, A. 18242 ; Кинтана, М. и др. 165 ; Розенгуртт, Б. В-5883.
Венесуэла: Буркарт, А.26783 .
Axonopus suffultus (J.
Leave a Comment