Колоски как нарисовать: Как нарисовать пшеницу карандашом поэтапно
18.04.2021 


 Разное
Разное
Рисование в старшей группе на тему: «Натюрморт с колосками» | План-конспект занятия по рисованию (старшая группа):
Приоритетная образовательная область: художественно-эстетическое развитие (рисование)
Интеграция с другими образовательными областями: познавательное развитие, физическое развитие, речевое развитие.
Развивающая предметно-пространственная среда темы НОД: в групповой комнате; гуашь, кисточки, стаканчики с водой, бумага размером альбомного листа (на каждого ребенка) ; игрушка Петушок, аудиозапись пения петушка.
Цель: Формировать умение детей рисовать концом кисти (тонкий стебель, закреплять навык примакивания, знания цветов и оттенков, изображать вазу простой формы, аккуратно закрашивать ее форму, развивать чувство композиции, технические навыки, воспитывать чувство уважения к труду взрослых, трудолюбие, аккуратность.
Содержание НОД:
Включаю звук пения Петушка.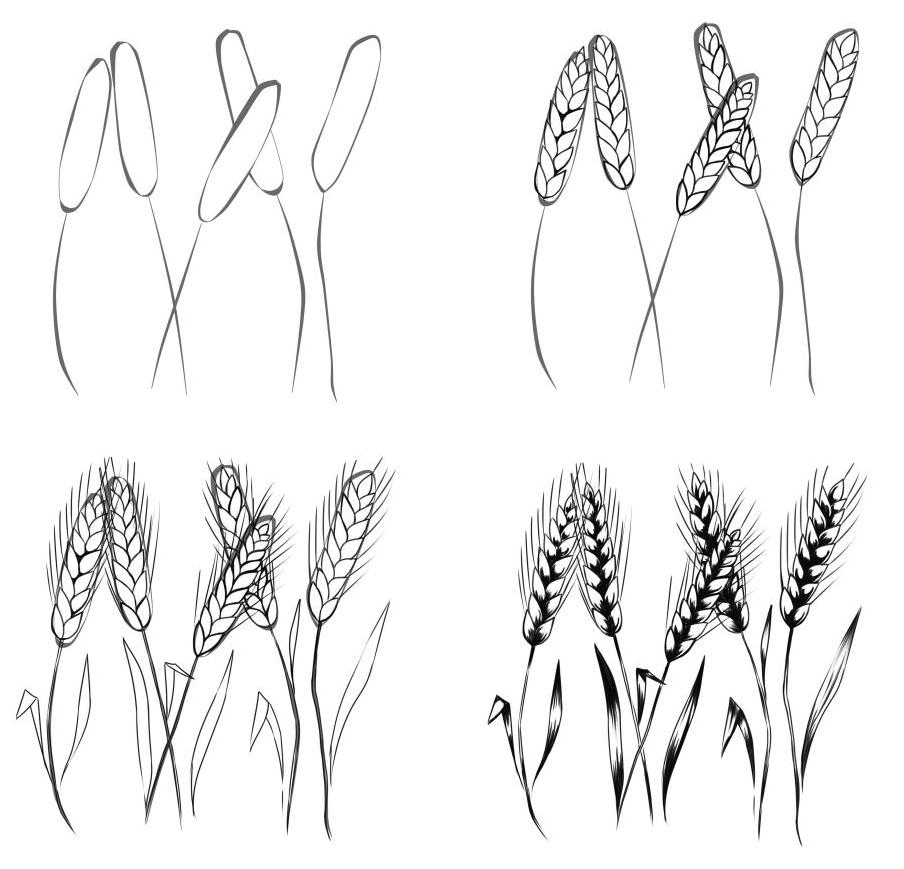
— Ой, ребята, это кто? Где же он прячется (дети находят)
— Как вы думаете, почему он спрятался?
— Петушок спрятался, ребята, потому что расстроился. Он нашел только один колосок. Дело в том, что у Петушка скоро день рождения и ему нужны колоски, чтобы из них сделать муку и спечь каравай.
— А может Петушок, из одного колоска сделать каравай?
— Почему? (В одном колоске мало зернышек, а значит получиться мало муки)
— Как же помочь Петушку?
— А как мы можем сделать много колосков? (вылепить, нарисовать, вырезать)
— Посмотрите на стол, и вы узнаете, чем мы будем работать и что делать (рисовать)
Давайте мы сегодня с вами сделаем много колосков. Вот посмотрите, какие они, какая форма у зернышек, какой цвет, как они расположены, их размер?
(Дети рассматривают колоски, трогают ручками, и отмечают: форма зернышек овальная; цвет – желтый, светло коричневый; расположены — один за другим; размер – маленькие)
— Каждый из вас соберет свои колоски в вазу и подарит Петушку, согласны?
— Я покажу вам, как можно нарисовать колоски в вазе. Сначала я нарисую вазу. Какой формы бывают вазы? (круглые, прямоугольные, овальные, низкие, высокие). Ребята, обратите внимание, что ваза должна быть до середины листа, а иначе колоски у вас не поместятся.
Сначала я нарисую вазу. Какой формы бывают вазы? (круглые, прямоугольные, овальные, низкие, высокие). Ребята, обратите внимание, что ваза должна быть до середины листа, а иначе колоски у вас не поместятся.
-Берем большую кисточку в руку, окунаем хвостик кисточки в воду, лишнюю капельку отжимаем о край стаканчика, набираем гуашь на хвостик, окунаем только хвостик, чтобы кисточка не испачкалась. Рисуем стебель снизу вверх, находим середину стебля и ставим точку, от этой точки вверх по разные стороны стебелька рисуем кармашки, в которых находятся зёрнышки техникой «примакивание». Вспоминаем, как мы это делаем. Прикладываем хвостик кисточки к бумаге и сразу же убираем. Получается ровный, красивый отпечаток. И последний раз примакнем кисточкой на самом верху колоска. Теперь берём тонкую кисточку и рисуем усики у колоска, аккуратно между кармашками. Они называются остья. А кто уже нарисовал остья, можно нарисовать листочки.
— Ребята, давайте поиграем с Вами, вспомним, что нужно, чтобы выросли колоски.
Физминутка «В землю зёрнышко попало»:
1. В землю зёрнышко попало. (Дети присаживаются на корточки, прижимают голову к коленям и закрывают её руками)
2. Прорастать на солнце стало (Дети поднимают руки вверх над головой, приговаривая) :
«Золотое солнышко, ты согрей нам зёрнышко!»
«Дождик, дождик, поливай! Будет хлеба урожай!» (Дети вытягивают руки вперёд, ладонями вверх и «брызгают» на себя водой.)
3. Дождик землю поливал, и росточек подрастал! (Дети медленно встают) .
4. К свету и теплу тянулся (руки в стороны)
5. И красавцем обернулся (руки вперёд) .
6. Золотые колоски на ветру качаются, (Дети встают на носочки, поднимают прямые руки вверх, кисти рук опущены вниз) .
7. Низко наклоняются (Дети машут поднятыми вверх руками и приговаривают) :
«Поспел урожай! Приезжай, убирай! (наклоны вниз)
-Давайте вспомним, как надо сидеть, когда рисуем: «Сяду прямо не согнусь, за работу я возьмусь!»
Воспитатель рисует на мольберте и комментирует.
Дети рисуют самостоятельно по образцу.
Петушок любуется работами детей, хвалит ребят и благодарит их за помощь!
— Вот теперь Петушку хватит и на каравай и на пироги.
Рефлексия:
О каком жанре искусства мы сегодня говорили? (натюрморт)
Что на занятии мы рисовали? (вазу с колосками)
Трудно было рисовать, что именно? (ответы детей)
Понравились вам ваши рисунки? Нравится вам заниматься рисованием? (ДА)
Сказка на ночь про колосок. Читать и слушать
Рос-рос колосок. Чтобы ему по-настоящему вытянуться, надо вдоволь насладиться солнышком, напиться доброй водицы. Что колосок и сделал. А когда он вырос, произошло вот что…
Слушать сказку (2мин54сек)
Сказка на ночь «Добрый колосок»
Автор: Ирис Ревю
Жил-был колосок. Он был небольшой, золотисто-желтого цвета.
— Кем же я стану, когда вырасту? – думал он. – Ватрушкой, пирожком или пышкой? Я ещё не определился, кем буду.
А колосок тем временем рос, наливался соком, улыбался золотому солнышку. И всё время думал о своем будущем.
— Хорошо бы стать сладкой булочкой, калачом или слойкой. А вот быть сушкой или сухарем мне не хотелось бы, ведь я такой нежный.
Колосок продолжал расти. Он был уже довольно большим и крепким. С ветром он подружился, дождик никогда не забывал поливать его, а солнышко посылало колоску самый весёлый свой лучик.
— Мне хотелось бы быть крендельком, лепешкой или пончиком. Тоже неплохо, — думал колосок.
Лето заканчивалось. Колосок превратился в большой, красивый колос. Его собрали, обратили в муку, а потом отправили к хлебопекарю.
Наш знакомый колосок и ещё сотни таких колосков стали большим, вкусным караваем. Ах, какой красивый был этот каравай!
Он попал на праздник к детям. У мальчика Василька, а попросту Васи, был день рождения. Дети водили хоровод и пели:
— Каравай, каравай, кого хочешь выбирай!
А потом принялись за настоящий, ароматный каравай.
Был на дне рождении Василька и необычный гость – старый сказочник. Он рассказывал детям сказки, в том числе и про колосок. Старый сказочник поведал, что весной в поле появятся новые колоски, из которых сделают ароматный хлеб.
Когда солнышко катилось к закату, дети стали расходится по домам. Василек долго думал про колосок, потом нарисовал его и лёг спать.
Ты тоже, дружок, можешь нарисовать колосок: сначала маленький, а потом большой, высокий. А сейчас пора в Царство счастливых снов.
Спокойной ночи!
Нарисовать колосья пшеницы. «Колосок»
Елена Баранова
«Колосок». Конспект НОД по ОО «Художественно-эстетическое развитие» (рисование) в старшей группе компенсирующего вида
Проектировочный компонент .
Тема : «Колосок » .
Интеграция образовательных областей : художественно-эстетическое развитие , физическое развитие , речевое развитие , познавательное развитие , социально-коммуникативное развитие .
Цель : обучение детей рисованию колоска нетрадиционной техникой «примакивание» .
Задачи :
Коррекционно-образовательные : расширить знания детей о колоске , учить рисовать колосок гуашевой краской , передавая особенности его строения, закрепить навыки рисования с помощью примакивания ворса кисточки, активировать в речи детей прилагательное (золотистый, существительное (остья) .
Коррекционно-воспитательные : воспитывать художественный вкус , воспитывать желание помогать другим,
Коррекционно-развивающие : развивать слуховое внимание , мышление, мелкую моторику, развивать чувство ритма , развивать эстетическое восприятие.
Оборудование и материалы :
Театр мягкой игрушки (петушок, мышата, солнышко, игрушечная избушка, игрушечный веник, натуральный пшеничный колосок , мольберт, альбомные листы формата А4, гуашь золотистая, кисти двух видов(№6 и №2, стаканчики с водой, салфетки (на каждого ребёнка, каравай.
Детская деятельность :
игровая – игра «Каравай» , физминутка с речевым сопровождением; коммуникативная — вопросы-ответы,игра «Каравай» ; изобразительная- рисование ; музыкальная- пение песни «Солнышко» , «Каравай» , музыкально-ритмические движения в физминутке «В землю солнышко попало» ; познавательно-исследовательская- рассматривание колоска , двигательная- овладение основными движениями в игре «Каравай» , восприятие художественной литературы и фольклора : слушание сказки.
Предварительная работа : чтение украинской сказки «Колосок » , рассматривание колосков , рассматривание иллюстраций, просмотр мультфильма
Организационный компонент :
Подготовка к занятию :
5 баночек с золотистой гуашью, 5 альбомных листов бумаги, 5 салфеток,10 кисточек, 5 стаканчиков с водой.
Проведение занятий в соответствии с СанПиНом 2.4.1.3049-13 и здоровьесберегающими технологиями.
Продолжительность непрерывной непосредственно образовательной деятельности — не более 25 минут, в середине времени, отведённого на НОД, проведение физкультминутки.
Метод :
1) Наглядный,
2) Словесный,
3) Практический,
4) Игровой.
Приемы : загадка, игра, показ, пояснение.
Мотивация :
Театр мягкой игрушки (петушок, мышата) .
Ход занятия:
Ребята, посмотрите к нам пришли гости. Давайте с ними поздороваемся.
Дети здороваются.
Можно нашим гостям поприсутствовать у нас.
Дети разрешают.
Сегодня я хочу рассказать Вам сказку. Хотите её послушать?
Дети соглашаются.
Хорошо. Тогда занимайте любые свободные места.
Жили-были два мышонка, Круть и Верть, да петушок Голосистое Горлышко. Мышата только и знают, что танцуют и играют, а петушок спозаранку встанет, всех песней разбудит.
Давайте, ребята, поможем нашему петушку разбудить солнышко.
Дети поют песенку и выполняют движения :
Солнышко, солнышко, (изображение руками солнышка)
Выгляни в окошко. (изображение руками окошка)
На ребяток погляди, (руки в стороны)
Ты ребяткам посвети! (изображение руками фонариков)
Выглядывает солнышко.
Петушок
Спасибо, ребята, за помощь! Пойду я во двор подметать.
Ой, что я нашёл! Но сначала, отгадайте загадку :
Золотист он и усат,
В ста карманах сто ребят.
Догадались?
Ответы детей.
Правильно, это колосок . Цвет у него золотистый. Вот усы, вот кармашки, а вот и ребята зернышки (показывает) . А ещё, что есть у колоска ?
Ответы детей.
Позову-ка я мышат.
Круть, Верть! Глядите, что я нашёл!
Это колосок ! Нужно его обмолотить!
А кто его обмолотит?
Только не я!
Только не я!
Ладно, я обмолочу!
Ребята, давайте поиграем с Вами, вспомним, что нужно, чтобы выросли колоски , а потом я обмолочу колосок .
Физкультурная минутка «В землю зёрнышко попало» (под музыку музыкального руководителя) :
1. В землю зёрнышко попало.
(Дети присаживаются на корточки, прижимают голову к коленям и закрывают её руками)
2. Прорастать на солнце стало.
(Дети поднимают руки вверх над головой, приговаривая :
«Золотое солнышко, ты согрей нам зёрнышко!»
«Дождик, дождик, поливай! Будет хлеба урожай!» )
(Дети вытягивают руки вперёд, ладонями вверх и «брызгают» на себя водой.)
3. Дождик землю поливал, и росточек подрастал!
(Дети медленно встают) .
4. К свету и теплу тянулся (руки в стороны)
5. И красавцем обернулся (руки вперёд) .
6. Золотые колоски на ветру качаются ,
(Дети встают на носочки, поднимают прямые руки вверх, кисти рук опущены вниз).
7. Низко наклоняются.
(Дети машут поднятыми вверх руками и приговаривают :
«Поспел урожай! Приезжай, убирай!
(Наклоны вниз)
Спасибо, ребята! А где мой колосок ? У меня сегодня именины и я хотел испечь каравай!
Воспитатель :
Не расстраивайся, петушок. Ребята, давайте поможем петушку, нарисуем ему колоски .
Ребята, давайте поможем петушку, нарисуем ему колоски .
Дети соглашаются и садятся за столы.
Давайте вспомним, как надо сидеть, когда рисуем : «Сяду прямо не согнусь, за работу я возьмусь!»
Воспитатель рисует на мольберте и комментирует.
Берем большую кисточку в руку, окунаем хвостик кисточки в воду, лишнюю капельку отжимаем о край стаканчика, набираем гуашь на хвостик, окунаем только хвостик, чтобы кисточка не испачкалась. Рисуем стебель снизу вверх, находим середину стебля и ставим точку, от этой точки вверх по разные стороны стебелька рисуем кармашки, в которых находятся зёрнышки техникой «примакивание» . Вспоминаем, как мы это делаем. Прикладываем хвостик кисточки к бумаге и сразу же убираем. Шлёп-шлёп-шлёп. Получается ровный, красивый отпечаток. И последний раз шлёпнем кисточкой на самом верху колоска . Теперь берём тонкую кисточку и рисуем усики у колоска , аккуратно между кармашками. Они называются остья. А кто уже нарисовал остья , можно нарисовать листочки .
Дети рисуют (самостоятельная деятельность детей, воспитатель помогает по мере необходимости индивидуально каждому ребёнку и хвалит их).
Ребята, положите свои рисунки на один стол! Пусть они высохнут!
Ребята, что мы с Вами сегодня делали?
Ответы детей.
Что есть у колоска ?
Ответы детей.
Как называются усики у колоска ?
Ответы детей.
Петушок любуется работами детей, хвалит ребят и благодарит их за помощь!
Сюрпризный момент :
Прибегают мышата и приносят каравай!
Петушок, поздравляем тебя с именинами! Это мы взяли твой колосок , обмолотили его, отнесли на мельницу, намололи муки, истопили печь, замесили тесто и испекли каравай для тебя!
Спасибо, мышата!
Ребята, давайте поздравим петушка и споём ему каравай!
Дети поздравляют петушка и поют каравай (под музыку музыкального руководителя) :
Как на Петины именины испекли мы каравай :
(дети водят хоровод вокруг петушка)
Вот такой вышины,
(поднимают сомкнутые руки вверх)
Вот такой нижины,
(дети присаживаясь на корточки, опускают руки)
Вот такой ширины.
(дети расходятся на ширину вытянутых рук, растягивая круг)
Каравай, каравай, кого любишь выбирай!
(петушок выбирает детей, обходя их по кругу)
Я, по правде, люблю всех, а вот (имя выбранного ребёнка) лучше всех!
(показывает на одного из детей в круге) .
Выбранный ребёнок и петушок танцуют в кругу. Игра повторяется, пока все дети не станцуют.
Петушок и мышата караваем угощают детей и прощаются с ними.
Рефлексивный компонент :
Цель и задачи были достигнуты, дети были активны, им всё понравилось. Продуктивный вид деятельности – рисование можно было заменить на лепку.
Литература :
1. Лыкова И. А Изобразительная деятельность в детском саду.
2. Украинская сказка «Колосок » .
3. Малова В. В. конспекты занятий по духовно- нравственному воспитанию дошкольников на материале русской народной культуры.
4. Аверина И. Е. Физкультурные минутки в детском саду.
Урок посвящается всем ботанам и обжорам. Вы увидите как рисовать пшеницу карандашом поэтапно. Естественно рисовать целое поле я не собираюсь, зато покажу несколько колосков:
Пшеница это съедобная трава. Из нее делают много вкусностей: хлеб, макарошки, сладости, пиво и другие благородные напитки. Так что ее любят и взрослые и дети. Правда, выращивать, молоть, и обрабатывать никто не любит, но это уже совсем другой разговор. Дабы изобразить колосок пщеницы надо много терпения и внимательности. Особой техники изображения здесь нету, тем более не нужно соблюдать пропорции. Видов и сортов пшеницы настолько много, что она сама даже не знает какой вырастет. С другой стороны надо сделать красиво, повторяйте за мной.
Как нарисовать пшеницу карандашом поэтапноШаг первый. Делаем набросок, в виде камыша.
Шаг второй. Добавляем кругленькие формы, похожие на вербу.
Шаг третий. Поправляем контуры, добавляем усики.
Задание 40
Задание 41
По вертикали:
1.![]() Мишин папа печет хлеб. Он (пекарь)
Мишин папа печет хлеб. Он (пекарь)
По горизонтали:
2. Из кадушки вылезло пышное (тесто)
3. На Пасху бабушка испекла сдобный (кулич)
4. Петя съел сладкий медовый (пряник)
Задание 42
Допиши текст, не повторяя слов.
В нашей семье все любят изделия из сдобного теста. Бабушка часто готовит шаньги.
К чаю мама подаёт на стол булочки и плюшки, а по праздникам торт. Я хожу в магазин и покупаю для дедушки слойку, для папы батон, для мамы круассаны, для себя ватрушку.
***Задание 43
Выскажите предположения: какие условия нужны были для того, чтобы люди занимались этой деятельностью?
1. Ловля рыбы: водоем (река, озеро, море)
2. Обработка дерева: большие лесные массивы.
3. Производство металла: месторождения руды и угля.
4. Изготовление посуды: залежи глины, кварцевого песка. (для глиняной и стеклянной посуды).
Задание 44
Заполни схему.
Задание 45
1. Вспашка поля.
Вспашка поля.
2. Уборка урожая.
3. Изготовление ткани (ткачество)
4. Пастьба животных
5. Плетение лаптей.
Задание 46
Найди правильный ответ.
2. Крепостное право было отменено в XIX (19) веке.
*** Задание 47
Сочини рассказ по картине Н.В. Неврева «Торг. Сцена из крепостного быта. Из недавнего прошлого» (история девушки. См. с.78 учебника)
План рассказа
1. кем была девушка? Как её звали?
2. Почему помещик продает ее?
3. Что девушка чувствует? Может ли она возразить хозяину?
4. О чем говорит обстановка комнаты в доме помещика.
На картине мы видим двух помещиков. Напыщенные, высокомерные, облысевшие, они обсуждают сделку купли – продажи. Помещики спокойны, для них это привычное дело. Один хочет продать подороже, а другой купить подешевле. Товаром сделки выступает молодая крестьянская девушка по имени Аграфена (Груня). Она искусная вышивальшица, а у соседа помещика портняжная мастерская. Отведя в сторону взор, владелец девушки о чем-то размышляет, вероятно обдумывает достаточна ли предложенная цена. Покупатель уже хозяйственно положил руку на крепостную, уверенный, что дело сладится.
Отведя в сторону взор, владелец девушки о чем-то размышляет, вероятно обдумывает достаточна ли предложенная цена. Покупатель уже хозяйственно положил руку на крепостную, уверенный, что дело сладится.
Лицо и поза продаваемой девушки, крепко сжатые губы, непроницаемый взгляд в купе со сложенными на груди руками, выражает одновременно и протест и осознание бессилия.
Толпа крепостных на заднем плане – это бесправная людская масса, которая только и может, что посочувствовать, каждый из них знает, что его тоже могут вот так же продать. Разве, что выделяется среди них староста, который пригнал девушку на торг. Подпоясанный кушаком, он стоит безмолвный, безучастный и бесстрастный – он исполнитель барской воли.
Управление общего образования
Ртищевского района Саратовской области
Муниципальное дошкольное образовательное учреждение
« Детский сад №12 « Звездочка». Г. Ртищево Саратовской области»
Конспект рисование
Тема;«Хлеб – всему голова»
Воспитатель: Коломиец В. Э.
Э.
2016
Конспект рисование «Хлеб – всему голова»
Цель: Обучить детей нетрадиционным способам рисования пшеничного колоса.
Расширить и углубить знание детей о пшеничном колосе, его внешнем виде.
закрепить навыки рисования с помощью ворса кисточки.
— повышать мотивацию изобразительной деятельности через осознание ее нравственной значимости; совершенствовать общую и мелкую моторику.
Активизировать в речи детей прилагательные (желтый, колючий, золотистый).
Воспитательные задачи: воспитывать самостоятельность и целенаправленность в работе, умение доводить начатое дело до конца, аккуратность при работе с красками. Воспитывать бережное отношение к хлебу.
Ход занятия
Воспитатель рассказывает древнюю легенду о хлебном колосе:
В далекие времена, когда сам Бог ходил по земле, жизнь людей была легкой и сытой. Круглый год стояло лето. Когда нужно шел дождик, когда нужно – светило солнце. В лесах росли плодовые деревья. Звери были смирными, и все кормились травой. Люди не знали ни болезней, ни бед, ни голода. Хлебный колос был очень большим – стебля почти не было, зерна начинались от самой земли, и каждое зерно было размером с боб. Хлеба было так много, что его никто не ценил. Один раз Бог ходил по земле и увидел, как мать подтерла краюшкой только что испеченного хлеба обмаравшегося ребенка и при этом отказала в пище страннику. Бог рассердился, поднялся на небо и лишил людей хлеба. Стала земля как камень, реки высохли, трава завяла. Наступил голод. Тогда кошка и собака пошли к Богу просить хлеба. Тот сжалился и выделил на кошачью и собачью долю маленький колос на длинном стебле. Бог сделал так, что лето стало занимать только половину года. Зима – для людей, а лето – для зверей. Люди, выпекая свежий хлеб, первый кусок отдавали кошкам и собакам. А теперь скажите, чему учит легенда?
Люди не знали ни болезней, ни бед, ни голода. Хлебный колос был очень большим – стебля почти не было, зерна начинались от самой земли, и каждое зерно было размером с боб. Хлеба было так много, что его никто не ценил. Один раз Бог ходил по земле и увидел, как мать подтерла краюшкой только что испеченного хлеба обмаравшегося ребенка и при этом отказала в пище страннику. Бог рассердился, поднялся на небо и лишил людей хлеба. Стала земля как камень, реки высохли, трава завяла. Наступил голод. Тогда кошка и собака пошли к Богу просить хлеба. Тот сжалился и выделил на кошачью и собачью долю маленький колос на длинном стебле. Бог сделал так, что лето стало занимать только половину года. Зима – для людей, а лето – для зверей. Люди, выпекая свежий хлеб, первый кусок отдавали кошкам и собакам. А теперь скажите, чему учит легенда?
Воспитатель:
ребята, отгадайте загадку, «вырос в поле дом, полон дом зерном» (колос). Вот колосок у нас сегодня в гостях, давайте расскажем, что мы про него знаем. Где растет колосок? Какие растения еще растут в поле, у которых есть колоски?
Где растет колосок? Какие растения еще растут в поле, у которых есть колоски?
Дети: пшеница, рожь, ячмень.
Воспитатель: ребята, давайте посмотрим на картинку, где изображено поле пшеницы. Что находится на колоске?
Дети: зерна (воспитатель демонстрирует натуральный пшеничный колос)
Физкультминутка
В землю зернышко попало (дети приседают),
Прорастать на солнце стало (руки над головой).
Дождик землю поливал, и росточек подрастал (дети медленно встают).
К свету и теплу тянулся, и красавцем обернулся.
Воспитатель: дети, давайте присядем за стол и попробуем нарисовать пшеничное поле (в начале рисуем стебелек, а затем промакиванием ворса кисточки.
Воспитатель просит детей представить, что он – колосок, а они – зернышки. Дети медленно собираются в кружок на ковре, в центре находится воспитатель.
Ребята, скажите, что делают из пшеничных зерен? (муку разных сортов)
Урок посвящается всем ботанам и обжорам. Вы увидите как рисовать пшеницу карандашом поэтапно. Естественно рисовать целое я не собираюсь, зато покажу несколько колосков:
Естественно рисовать целое я не собираюсь, зато покажу несколько колосков:
Пшеница это съедобная трава. Из нее делают много вкусностей: хлеб, макарошки, сладости, пиво и другие благородные напитки. Так что ее любят и взрослые и дети. Правда, выращивать, молоть, и обрабатывать никто не любит, но это уже совсем другой разговор. Дабы изобразить колосок пщеницы надо много терпения и внимательности. Особой техники изображения здесь нету, тем более не нужно соблюдать пропорции. Видов и сортов пшеницы настолько много, что она сама даже не знает какой вырастет. С другой стороны надо сделать красиво, повторяйте за мной.
Как нарисовать пшеницу карандашом поэтапноШаг первый. Делаем набросок, в виде камыша. Шаг второй. Добавляем кругленькие формы, похожие на . Шаг третий. Поправляем контуры, добавляем усики. Шаг четвертый. Добавим штриховки и большие усики. Готово: Попробуйте изобразить еще такие растения.
Как нарисовать пшеницу, колосья, поэтапно?
Камут, или пшеница Хорасан (правильное название), была найдена в гробницах фараонов в 1949 году. Возможно именно по этому ее и называют «Зерно Фараонов».
Возможно именно по этому ее и называют «Зерно Фараонов».
зерно пшеницы отличается от нашего размером, и содержанием полезных веществ. Для примера, в Хорасан на 30 процентов больше содержание белков и клетчатки. Кроме этого, зерно богато на магний, цинк, селен, марганец и жирные кислоты.
Питательные свойства зерна превосходят в несколько раз характеристики привычной нам пшеницы, и именно по этому Камут пользуется популярностью во всем мире.
Собранное зерно уже достаточно твердое, но можно еще подсушить. По твердости будет напоминать камень.
После этого колосья бьют (берут сноп и лупят по бревну или полу так, чтобы зерна отлетали — желательно соответствующее помещение). Часть зерен может остаться, их можно выбить отдельно или отделить руками, крутя-перекатывая колосья между ладоней.
На зернах может остаться шелуха, для чего их надо катать, провеивать (подкидывать, чтобы легкую шелуху сдуло, а твердые тяжелые зерна сразу падали вниз). Ну и т.д.
Ну и т.д.
Самоопылением. Пыльца с тычинок переносится на пестик. (А если быть точнее,то у пшеницы облегатный способ опыления)
Ну а мы как понимаем? На запах, на вкус, смотрим как другие делают.
Т.е. если пахнет едой, значит еда и есть.
Если на вкус норм — тоже ок.
А если все стадо жует — тоже жуй, че сложного?
Пищевая ценность пророщенных зерен пшеницы значительно выше,чем в сухих зернах,так как формирующийся росток под влиянием химических процессов,меняет качественный состав.Зерно обогащается витаминами,микро-микроэлементами после прорастания,причем содержание витаминов и других полезных веществ существенно удваивается.Поэтому пищевая ценность пророщенных зерен пшеницы огромна.Употребляя такую еду,налаживаются все внутренние органы человека. Повышается сопротивляемость организма к инфекциям,поднимается иммунитет,нормализуется обмен веществ.То есть это живая еда для питания человека,которая наполняет тело энергией и силой.Особенно полезно питаться такой едой в зимне-весенний период,когда не хватает витаминов в организме человека.
Повышается сопротивляемость организма к инфекциям,поднимается иммунитет,нормализуется обмен веществ.То есть это живая еда для питания человека,которая наполняет тело энергией и силой.Особенно полезно питаться такой едой в зимне-весенний период,когда не хватает витаминов в организме человека.
Рисование и живопись — Вальдорфский садик Колосок
Одним из тех занятий, в которых участвует вся группа, является рисование. При этом речь идёт не о том, как заставить детей рисовать нечто определённое, например, дом, дерево и т.д. Ребёнок испытывает естественную потребность прежде всего просто повозиться с красками как таковыми.
Дети наносят акварель на бумагу широкой кистью и живут вместе с ложащимися на лист красками. Они радуются их сгущению и размытости, тому, что из смеси красок то там, то сям возникает новый цвет.
Живопись по-мокрому акварельными красками — по-настоящему волшебное и завораживающее занятие. Задача педагогов в этом процессе — дать возможность играть с цветом, обеспечить его восприятие во всей полноте.
Дети не спрашивают, что им рисовать, они сразу принимаются за дело. Если бы тема рисунка была задана, пришлось бы апеллировать к представлениям и памяти. Некоторые довольно долго рисуют одним цветом, прежде чем добавить второй или третий. Другие рисуют сразу несколькими красками. После рисования работы не обсуждаются. И воспитатель радуется вместе с детьми, вместе с ними он наблюдает за тем, что могут краски.
Существует три цвета, которые нельзя получить, смешивая другие цвета. Это три основных цвета: жёлтый, синий и красный. Многообразие остальных цветов можно получить, смешивая три основных цвета в разных пропорциях. В вальдорфском детском саду мы не обучаем детей теории цвета, но предоставляем им возможность пережить возникновение различных цветов на бумаге и творчески переработать пережитое.
Смешение акварельных жидких красок на мокрой бумаге даёт детям живую подвижную картину.
Никаких оценок рисункам мы не даём, ведь дети рисуют красиво, они все — художники. Высохшие рисунки раскладываются на стеллаж по именам детей и хранятся год. В конце года собираем их в виде альбомов и дарим детям.
Высохшие рисунки раскладываются на стеллаж по именам детей и хранятся год. В конце года собираем их в виде альбомов и дарим детям.
Радуги мостик весь расписной
Рисование, живопись — вот та деятельность, где дети не должны подражать.
В рисунках ребёнок отражает свое внутреннее состояние, поэтому на рисунок ребёнка не должен влиять взрослый или кто-либо другой и не надо учить ребёнка «рисовать правильно».
По детским рисункам, по их цветовому решению можно увидеть развитие, состояние детей и даже их болезни.
Мы не даём детям никаких тем или указаний для рисования. Они рисуют не карандашами, а цветными мелками прямоугольной формы, чтобы можно было рисовать плоскостями. Рисунок восковыми мелками даёт более подвижную картинку, что соответствует образному мышлению ребёнка. В этих как бы размытых рисунках он сам видит свои образы, и надо дать ему в этом полную свободу. Цветные, а тем более простые карандаши приближают ребёнка к графике, а чёткие графические линии несут в себе жёсткость, некоторую завершенность и тем ограничивают его фантазию. После семи лет ребёнку можно предлагать цветные карандаши.
После семи лет ребёнку можно предлагать цветные карандаши.
Для развития художественных способностей ребёнка важно переживание цвета. Поэтому раз в неделю идут занятия живописью, акварельными красками. Дети рисуют жидкими красками по мокрой бумаге тремя основными цветами. Сначала начинаем рисовать только одним цветом (после лета это может быть жёлтый) целый месяц, и дети переживают этот цвет. Затем даём второй цвет — красный. Жёлтая краска разливается по мокрой бумаге, встречается с красной. А глазах у ребёнка рождается новый цвет — оранжевый — в разных его оттенках. Это чудо! Это радость, которую дети активно переживают.
В другие дни дети рисуют светло-красным цветом, розовым. Потом только синим, начиная с самых светлых оттенков. Это бывает в конце осени — начале зимы. Синий цвет, все его оттенки, дети видят в зимнем утреннем небе.
Смешанные цвета рождаются у ребёнка на глазах. Это самостоятельное открытие мира цветов и оттенков очень важно для ребёнка.
Старшим детям можно давать сразу три цвета. Работа с тремя основными цветами развивает у ребёнка чувство цвета, будит фантазию в создании и переживании гаммы оттенков. Правильное воспитание ребёнка к восприятию разных цветов как в живописи, так и в жизни — залог гармоничного развития человека.
Нарисовать хлебный злак поэтапно. «Колосок». Конспект НОД по ОО «Художественно-эстетическое развитие» (рисование) в старшей группе компенсирующего вида
Управление общего образования
Ртищевского района Саратовской области
Муниципальное дошкольное образовательное учреждение
« Детский сад №12 « Звездочка». Г. Ртищево Саратовской области»
Конспект рисование
Тема;«Хлеб – всему голова»
Воспитатель: Коломиец В.Э.
2016
Конспект рисование «Хлеб – всему голова»
Цель: Обучить детей нетрадиционным способам рисования пшеничного колоса.
Расширить и углубить знание детей о пшеничном колосе, его внешнем виде.
закрепить навыки рисования с помощью ворса кисточки.
— повышать мотивацию изобразительной деятельности через осознание ее нравственной значимости; совершенствовать общую и мелкую моторику.
Активизировать в речи детей прилагательные (желтый, колючий, золотистый).
Воспитательные задачи: воспитывать самостоятельность и целенаправленность в работе, умение доводить начатое дело до конца, аккуратность при работе с красками. Воспитывать бережное отношение к хлебу.
Ход занятия
Воспитатель рассказывает древнюю легенду о хлебном колосе:
В далекие времена, когда сам Бог ходил по земле, жизнь людей была легкой и сытой. Круглый год стояло лето. Когда нужно шел дождик, когда нужно – светило солнце. В лесах росли плодовые деревья. Звери были смирными, и все кормились травой. Люди не знали ни болезней, ни бед, ни голода. Хлебный колос был очень большим – стебля почти не было, зерна начинались от самой земли, и каждое зерно было размером с боб. Хлеба было так много, что его никто не ценил. Один раз Бог ходил по земле и увидел, как мать подтерла краюшкой только что испеченного хлеба обмаравшегося ребенка и при этом отказала в пище страннику. Бог рассердился, поднялся на небо и лишил людей хлеба. Стала земля как камень, реки высохли, трава завяла. Наступил голод. Тогда кошка и собака пошли к Богу просить хлеба. Тот сжалился и выделил на кошачью и собачью долю маленький колос на длинном стебле. Бог сделал так, что лето стало занимать только половину года. Зима – для людей, а лето – для зверей. Люди, выпекая свежий хлеб, первый кусок отдавали кошкам и собакам. А теперь скажите, чему учит легенда?
Бог рассердился, поднялся на небо и лишил людей хлеба. Стала земля как камень, реки высохли, трава завяла. Наступил голод. Тогда кошка и собака пошли к Богу просить хлеба. Тот сжалился и выделил на кошачью и собачью долю маленький колос на длинном стебле. Бог сделал так, что лето стало занимать только половину года. Зима – для людей, а лето – для зверей. Люди, выпекая свежий хлеб, первый кусок отдавали кошкам и собакам. А теперь скажите, чему учит легенда?
Воспитатель: ребята, отгадайте загадку, «вырос в поле дом, полон дом зерном» (колос). Вот колосок у нас сегодня в гостях, давайте расскажем, что мы про него знаем. Где растет колосок? Какие растения еще растут в поле, у которых есть колоски?
Дети: пшеница, рожь, ячмень.
Воспитатель: ребята, давайте посмотрим на картинку, где изображено поле пшеницы. Что находится на колоске?
Дети: зерна (воспитатель демонстрирует натуральный пшеничный колос)
Физкультминутка
В землю зернышко попало (дети приседают),
Прорастать на солнце стало (руки над головой).
Дождик землю поливал, и росточек подрастал (дети медленно встают).
К свету и теплу тянулся, и красавцем обернулся.
Воспитатель: дети, давайте присядем за стол и попробуем нарисовать пшеничное поле (в начале рисуем стебелек, а затем промакиванием ворса кисточки.
Воспитатель просит детей представить, что он – колосок, а они – зернышки. Дети медленно собираются в кружок на ковре, в центре находится воспитатель.
Ребята, скажите, что делают из пшеничных зерен? (муку разных сортов)
Пшеница (лат. Triticum) – это одно из древнейших злаковых растений отдела цветковые, класса однодольные, порядка злакоцветные, семейства злаки.
Описание пшеницы и фотографии.
Все сорта пшеницы имеют основные характерные признаки. Высота стебля пшеницы достигает 30-150 сантиметров. Сами стебли полые и прямостоячие, с хорошо заметными узлами. С одного растения, как правило, вырастает до 12 стеблей. Листья пшеницы достигают ширины 20 мм, по форме они плоские и чаще всего линейные, с параллельными прожилками, волокнистые, на ощупь шершавые. Листовые влагалища пшеницы ярко выражены и хорошо развиты. Расщепленные до самого основания влагалища имеют на верхушке ланцетные ушки. Их язычки голые и перепончатые, от 0,5 до 3 мм длиной. Растение пшеница имеет мочковатую корневую систему.
Листовые влагалища пшеницы ярко выражены и хорошо развиты. Расщепленные до самого основания влагалища имеют на верхушке ланцетные ушки. Их язычки голые и перепончатые, от 0,5 до 3 мм длиной. Растение пшеница имеет мочковатую корневую систему.
Строение пшеницы, колосья.
Соцветие пшеницы – это прямой, сложный колос от 4 до 15 см длиной, бывает продолговатым или яйцевидным. На оси каждого колоса расположены колосовые чешуйки длиной 6-15 мм. Колосья пшеницы одиночные и примыкают к оси двумя одинаковыми рядами длиной 5-18 миллиметров, с несколькими сближенными цветками, которых чаще всего от 2 до 7. Ось колоса пшеницы не содержит сочленений. Цветок пшеницы имеет 2 чешуи и 2 пленки, 3 тычинки, пестик и 2 рыльца. Это строение является типичным для цветков злаковых растений. Когда созревает пшеница, она дает зерновые плоды.
Сорта и виды пшеницы.
Разновидностей пшеницы очень много. Эти растения имеют достаточно сложную классификацию, включающую в себя секции, виды и подвиды, а также около 10 гибридов, как внутриродовых, так и межродовых. Выделяют следующие виды пшеницы:
Выделяют следующие виды пшеницы:
- однолетние
- двухлетние
Пшеница яровая и озимая – отличия.
По сроку посева выделяется:
- Яровая пшеница – сеется с марта по май, созревает в течение 100 безморозных дней, убирают ее в начале осени. Более засухоустойчивая, чем озимая пшеница, обладает отличными хлебопекарными свойствами.
- Озимая пшеница – сеется в конце лета до середины осени, дает урожай в начале-середине лета следующего года. Дает более высокий урожай, но предпочитает районы с мягким климатом и снежной зимой.
Пшеница мягкая и твердая.
Виды пшеницы по твердости зерна:
- мягкая пшеница – имеет более широкий и короткий колос и более короткую или отсутствующую ость. Этот тип отличается высоким содержанием белка и клейковины. Из мягкой пшеницы делают муку.
- мягкая яровая краснозерная пшеница – к данному типу относятся сорта пшеницы Алтайская 81, Воронежская 10, Люба, Московская 35 и т.
 д.
д. - мягкая яровая белозерная пшеница – к данному типу относятся сорта пшеницы Новосибирская 67, Саратовская 55 и т.д.
- мягкая озимая краснозерная пшеница – к данному типу относятся сорта Донская безостая, Обрий, Волгоградская 84, Юна и т.д.
- мягкая озимая белозерная пшеница – к данному типу относятся сорта Кинсовская 3, Альбидум 28 и т.д.
- мягкая яровая краснозерная пшеница – к данному типу относятся сорта пшеницы Алтайская 81, Воронежская 10, Люба, Московская 35 и т.
- твердая пшеница – имеет колоски, более туго облегаемые наружными пленками, зерна из них не осыпаются, но их труднее выделить. Обладает насыщенным желтым цветом и приятным запахом. Твердая пшеница применяется для изготовления макаронных изделий.
- Пшеница твердая яровая (дурум) – к данному типу относятся сорта Алмаз, Оренбургская 2, Светлана и т.д.
- пшеница твердая озимая – к данному типу относятся сорта Вахт, Муганс, Парус и т.д.
Где растет пшеница?
Пшеница растет везде, кроме тропиков, поскольку многообразие специально созданных сортов позволяет использовать любые почвенные и климатические условия. Жара растению не страшна, если при этом отсутствует повышенная влажность, способствующая развитию болезней. Пшеница – растение настолько холодостойкое, что ее превосходит только ячмень и . Мягкая пшеница предпочитает влажный климат и распространена в Западной Европе, России Австралии. Твердая пшеница любит более сухой климат, ее выращивают в США, Канаде, Северной Африке, Азии. Озимая пшеница преобладает в тех районах, где ее не повреждают морозы, к примеру, на Северном Кавказе, в Центрально-Черноземном районе России. Яровая пшеница выращивается на Южном Урале, в Западной Сибири, на Алтае.
Жара растению не страшна, если при этом отсутствует повышенная влажность, способствующая развитию болезней. Пшеница – растение настолько холодостойкое, что ее превосходит только ячмень и . Мягкая пшеница предпочитает влажный климат и распространена в Западной Европе, России Австралии. Твердая пшеница любит более сухой климат, ее выращивают в США, Канаде, Северной Африке, Азии. Озимая пшеница преобладает в тех районах, где ее не повреждают морозы, к примеру, на Северном Кавказе, в Центрально-Черноземном районе России. Яровая пшеница выращивается на Южном Урале, в Западной Сибири, на Алтае.
Рожь и пшеница – отличия.
Рожь и пшеница – одни из самых популярных и незаменимых злаковых культур. Эти злаки имеют внешнюю схожесть, но также и много различий.
- Сорта пшеницы гораздо более разнообразны, чем сорта ржи.
- Пшеница имеет более широкое применение, чем рожь.
- Зерна имеют различный внешний вид и химический состав.
- Пшеница предъявляет больше требований к почве и климату, чем .

Выращивание пшеницы.
Высокая урожайность пшеницы достигается при правильной подготовке к ее посеву. Поле для пшеницы обрабатывают культиваторами и выравнивают поверхность для обеспечения хорошего контакта семян пшеницы с почвой и получения одновременных всходов. Посев пшеницы производится на глубину 3-5 см с междурядьями 15 см.
Пшеница – растение весьма влагозависимое, и поэтому хороший урожай требует регулярного полива. Для сухого климата больше подойдут твердые сорта пшеницы, они менее прихотливы в плане влаги. Рост пшеницы обеспечивается внесением удобрений. Посеянная пшеница убирается комбайном при полной зрелости зерна.
Как прорастить зерна пшеницы?
Прорастить зерна пшеницы в домашних условиях очень просто. Зерно необходимо поместить в стеклянную банку объемом 1 литр. Оно должно занимать не более 1/4-1/3 банки. В банку долить воду почти до краев, замочить зерна на 7-8 часов. После этого слить воду через марлю, промыть пшеницу и залить свежую воду на 3-4 часа. Таким образом, зерна пшеницы необходимо промывать 2-4 раза в день, давать воде стечь и затем снова помещать зерна в банку. Через сутки проростки достигнут высоты 1-2 мм, и проросшие зерна пшеницы уже можно будет есть.
Таким образом, зерна пшеницы необходимо промывать 2-4 раза в день, давать воде стечь и затем снова помещать зерна в банку. Через сутки проростки достигнут высоты 1-2 мм, и проросшие зерна пшеницы уже можно будет есть.
Как вырастить пшеницу в домашних условиях?
Зеленые ростки пшеницы можно получить, продолжив замачивать зерна еще 1-2 дня. Проростки размером 1-2 см необходимо пересадить в емкость с землей. Пророщенные зерна пшеницы укладывают на землю и засыпают сверху слоем земли в 1 см. Землю необходимо поливать, но не слишком обильно. Проростки пшеницы готовы к употреблению через несколько дней.
Пшеница — это однолетнее злаковое растение, которое выращивается в большинстве стран мира. Из нее делают муку, крупы, макаронные и кондитерские изделия, даже пиво, и это далеко не полный список. А нарисовать пшеницу совсем несложно. Ниже мы рассмотрим способы, как это сделать.
Как нарисовать пшеницу карандашом: материалы
Прежде чем приступать к рисованию пшеницы, подготовьте необходимые материалы. Это простые карандаши (лучше всего взять один твердый и один мягкий), лист бумаги, ластик и цветные карандаши (желтый, светло-оранжевый, зеленый, коричневый), если вы хотите раскрасить рисунок.
Это простые карандаши (лучше всего взять один твердый и один мягкий), лист бумаги, ластик и цветные карандаши (желтый, светло-оранжевый, зеленый, коричневый), если вы хотите раскрасить рисунок.
Первый способ
Сначала рассмотрим один из самых простых методов нарисовать пшеницу поэтапно, который подойдет для рисования с ребенком.
- На листе бумаги рисуем простым карандашом несколько слегка изогнутых вертикальных линий. Их количество зависит от того, сколько вы хотите нарисовать колосков.
- Вокруг верхней части одного из стеблей нарисуйте девять кружочков.
- Теперь сделайте из кругов зерна, которые похожи на капли по форме. Таким же образом нарисуйте зерна на остальных стеблях.
- На каждом колоске рисуем по несколько усиков. Их должно быть немного, по 4-5 штучек на каждом колоске. Снизу стебельков дорисовываем длинные и узкие листья.
Теперь вы знаете, как нарисовать пшеницу простым способом, можно попробовать более сложный метод. В конце пшеницу раскрасьте, используя оранжевые и желтые карандаши для колосков и зеленые для листьев.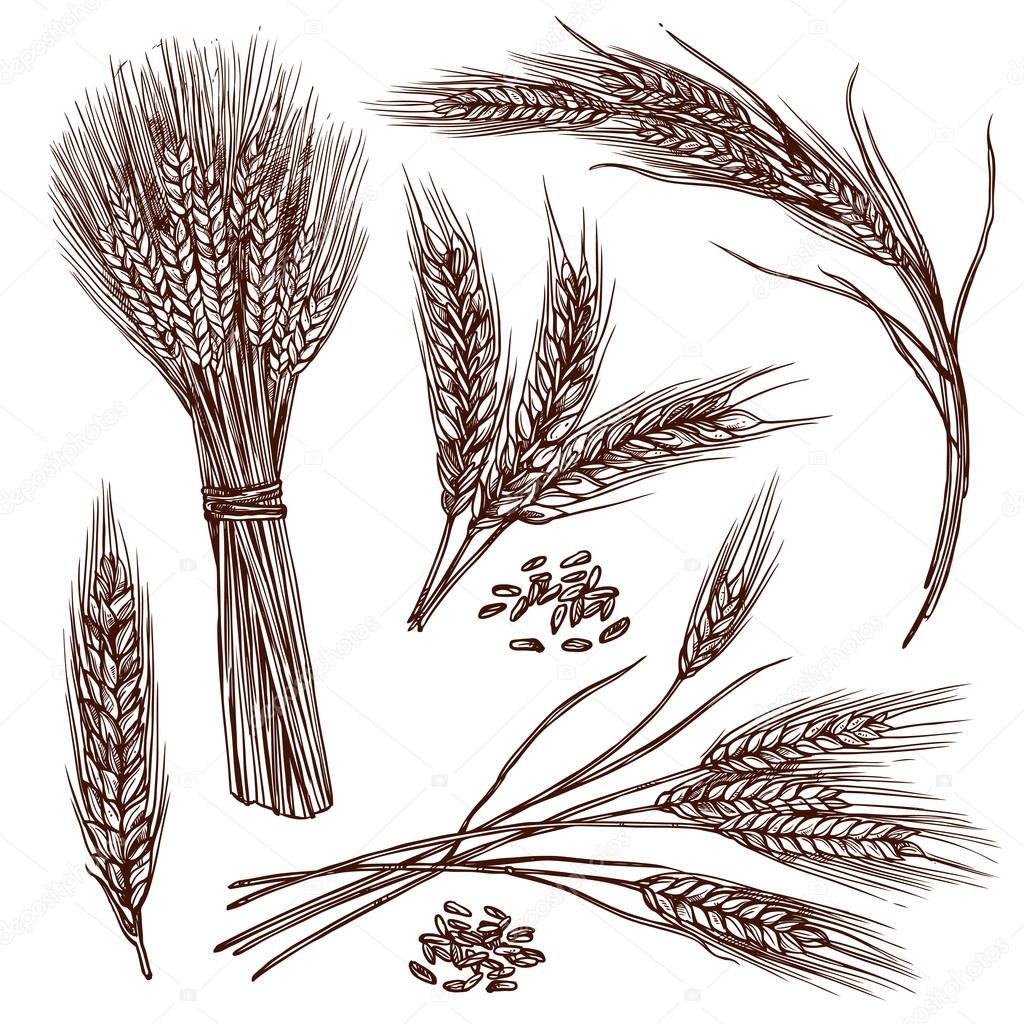
Второй способ
Как нарисовать пшеницу еще одним несложным способом? Для этого также потребуется лист бумаги, простой карандаш и ластик.
Рисуем тонкие, слегка наклоненные линии. В верхней части каждой линии изображаем колоски. Прорисовываем по несколько зернышек с каждой стороны и одно в самом верху. Зерна могут быть как в форме капелек, так и в виде маленьких овалов.
Проводим толстые полосы с двух сторон колоса, а сверху рисуем несколько коротких линий. Дорисовываем пару листочков рядом со стеблями. Вытираем ненужные линии — и рисунок готов. Можно дорисовать еще несколько колосков и раскрасить картинку с помощью красок или карандашей, используя желтый и коричневый цвет.
Третий способ
Если вы хотите знать, как нарисовать пшеницу чуть более реалистично, то вам подойдет этот метод.
Таким способом можно нарисовать как один колосок, так и целый сноп или даже поле. Кроме того, колоски можно немного видоизменить, разместив, например, зернышки на немного большем расстоянии друг от друга, или дорисовав сбоку дополнительный ряд зерен.![]()
Урок посвящается всем ботанам и обжорам. Вы увидите как рисовать пшеницу карандашом поэтапно. Естественно рисовать целое я не собираюсь, зато покажу несколько колосков:
Пшеница это съедобная трава. Из нее делают много вкусностей: хлеб, макарошки, сладости, пиво и другие благородные напитки. Так что ее любят и взрослые и дети. Правда, выращивать, молоть, и обрабатывать никто не любит, но это уже совсем другой разговор. Дабы изобразить колосок пщеницы надо много терпения и внимательности. Особой техники изображения здесь нету, тем более не нужно соблюдать пропорции. Видов и сортов пшеницы настолько много, что она сама даже не знает какой вырастет. С другой стороны надо сделать красиво, повторяйте за мной.
Как нарисовать пшеницу карандашом поэтапноШаг первый. Делаем набросок, в виде камыша. Шаг второй. Добавляем кругленькие формы, похожие на . Шаг третий. Поправляем контуры, добавляем усики. Шаг четвертый. Добавим штриховки и большие усики. Готово: Попробуйте изобразить еще такие растения.
Нетрадиционное рисование. Мастер-класс в 4 классе «Золотой колосок».
Панфилова Надежда Павловна, учитель начальных классов МБОУ «Раздольненская школа-гимназия №2 им Л.Рябики» Республика КрымМастер-класс пошагово «Золотой колосок». Нетрадиционная техника рисования для детей начальной школы.
Описание: мастер-класс будет интересен педагогам дополнительного образования, учителям начальной школы, воспитателям, творческим детям и родителям. Для человека хлеб- основная еда. Пшеница- самый главный хлеб. Название «пшеница» одинаково звучит на многих языках- пшеныця, вшениця. А само название её возникло от слова «пшоно»- «очищенное зерно».
«Уродись, пшеница,
С корня корениста,
Сверху колосиста.
Чтобы колосок
Был крепок, как дубок.
Чтобы зёрнышко
Было с вёдрышко!»
(Народная песня.)
Назначение: работа станет хорошим украшением интерьера, выставочным экспонатом, подарком.
Цель: создание рисунка нетрадиционным приёмом рисования.

Задачи:
-учить рисовать нетрадиционным приёмом- оттиском колоска;
-развивать воображение, творческие способности;
-развивать наблюдательность, эстетические чувства,
-воспитывать аккуратность, любовь к прекрасному, природе, уважение к людям труда.
Материалы:
-альбомный лист,
-краски, кисточки,
-вода, рамка,
-колоски пшеницы.
Хлебные растения, которыми питается человек, самые ценные. Это пшеница (озимая и яровая), рожь, ячмень,рис, кукуруза, просо, сорго и много других. Для человека хлеб- основная еда. Самый главный хлеб- пшеница,»ржи богатая сестрица». Учёные интересовались местом зарождения пшеницы. Русский учёный Николай Иванович Вавилов установил, что «прародиной» пшеницы является Азия, Закавказье. В горных районах и сейчас есть целые заросли дикого вида пшеницы.
Загадка.
Был крупинкой золотой,
Стал зелёною стрелой.
Солнце летнее светило,
И стрелу позолотило.
Что за стрела?
(Колос.
 )
) Солнце землю согревает,
Дождик щедро поливает,
К концу лета вышел срок-
Вырос в поле колосок.
Поле солнцем залитое,
Говорят, что золотое…
Колоски росли, трудились,
Светом солнечным налились,
Силу взяли от Земли-
Золотыми стать смогли!
(Светлана Богдан.)
Ход работы.
Альбомный лист кладём горизонтально. В верхней части листа наносим синим цветом узкую полоску.
Салфеткой убираем (промокаем) немного краски. Добавляем белую краску- промокаем.
Тёмно-зелёным цветом краски проводим линию горизонта.
Наносим жёлтым цветом горизонтальные линии.
Добавим оранжевой краски.
В нижней части листа добавим светло-коричневую краску.
На голубом фоне неба, на линии горизонта наметим очертания деревьев.
На коричневой полоске проводим вертикальные линии. Это стебли колосков.
На колосок пшеницы (надо обязательно отрезать усики) кисточкой наносим краску намного светлее коричневого фона.
 Аккуратно прижимаем колосок и делаем ряд отпечатков. Необходимо повторить несколько раз, и обязательно менять тон жёлтой краски.
Аккуратно прижимаем колосок и делаем ряд отпечатков. Необходимо повторить несколько раз, и обязательно менять тон жёлтой краски.В нижней части листа зелёным цветом наносим фон для травы.
В стеблях пшеницы растут васильки.
Работа «Золотые колоски»
Работы моих учеников.
Золотился в поле,
Спелый колосок
Из земли он черпал-
Самый свежий сок.
Умывался росами,
Небом любовался
И залётным ветром
Утром вытирался.
Он мужал и радовался,
В поле колосился и
С такими же как он
Дружно веселился.
(С.Богдан.)
МДОУ «Заволжский детский сад «Колосок». Советы специалистов
Как одевать ребенка на прогулку
Мамы постоянно борются с непреодолимым желанием одеть на ребенка как можно больше теплых вещей. Врачи часто с ужасом смотрят на детей, которые приходят на прием настолько укутанные, что рискуют не только вспотеть, но и получить тепловой удар в разгаре зимы. Объяснить такое явление несложно. Я уверен, что природа наделила каждого из нас несколько преувеличенным чувством беспокойства о своих отпрысках. Вероятно, существование этого инстинкта связано с жизненной необходимостью обеспечить своего ребенка в первую очередь теплом и едой, особенно тогда, когда он еще не может ничего сказать о своих желаниях. Молодые мамы, не имеющие опыта ухода за детьми, это беспокойство ощущают наиболее остро.
Объяснить такое явление несложно. Я уверен, что природа наделила каждого из нас несколько преувеличенным чувством беспокойства о своих отпрысках. Вероятно, существование этого инстинкта связано с жизненной необходимостью обеспечить своего ребенка в первую очередь теплом и едой, особенно тогда, когда он еще не может ничего сказать о своих желаниях. Молодые мамы, не имеющие опыта ухода за детьми, это беспокойство ощущают наиболее остро.
Существует и противоположная крайность. Иногда встречаются мамы, которые стремятся всем своим поведением показать свободу от предрассудков. Поэтому они воспитывают ребенка «в строгости». Такие мамы одевают детей настолько легко, что приводят в ужас бабушек, дедушек и соседей.
Чтобы знать, как одеть ребенка, надо учитывать его возраст и состояние здоровья. Руки и ноги малыша в норме должны быть немного прохладными. Маме советую это знать и не спешить, одевать его теплее. Детей постарше советую приучать к тому, чтобы они сами заботились о себе и, соответственно, об одежде в ту или иную погоду. Безусловно, нельзя зимой одевать на ребенка только комбинезон на фланелевой подкладке, так как вполне очевидно, что он не обеспечивает достаточного тепла. Такой же комбинезон, только на хорошей теплой подкладке — одежда более удобная для детей, чем сковывающие движения ребенка пальто и отдельные брюки.
Безусловно, нельзя зимой одевать на ребенка только комбинезон на фланелевой подкладке, так как вполне очевидно, что он не обеспечивает достаточного тепла. Такой же комбинезон, только на хорошей теплой подкладке — одежда более удобная для детей, чем сковывающие движения ребенка пальто и отдельные брюки.
Иногда я с содроганием смотрю зимой на ребенка, одетого совсем в легкую одежду, которая годится лишь для теплой погоды, или, наоборот, на детей, одетых летом, и жаркое время дня, в шерстяные кофточки и байковые чепчики. Все в нашей жизни должно быть в меру!
Если дети балуются
Этот тип поведения довольно распространен среди младших дошкольников, особенно среди четырехлетних. Если дети дурачатся слишком часто или в неподходящее время, то это может вызвать серьезные проблемы.
Как предотвратить проблему
Предоставляйте детям больше возможностей баловаться и даже поощряйте их к этому в специально отведенное время. Если они будут дурачиться именно в это время, скажите им, что они поступают хорошо, потому что никому не мешают, взрослым весело на них смотреть. Иногда сами предлагайте побаловаться, помогая детям понять, что это вовсе не плохое поведение, если никому не мешаешь и не портишь вещи.
Иногда сами предлагайте побаловаться, помогая детям понять, что это вовсе не плохое поведение, если никому не мешаешь и не портишь вещи.
Как справиться с проблемой, если она уже есть
Может оказаться так, что ребенок, который слишком много дурачится или строит из себя шута, на самом деле пытается уйти от каких-то серьезных проблем в своей жизни. Он прячется под «маску», пытаясь избежать непосильного напряжения. Такого же рода поведение может развиться и вследствие того, что он видит внимание, восхищение других детей и некоторых взрослых.
Обсудите вместе с другими членами семьи и воспитателями, что может скрывать ребенок за «маской шута», что пытается заглушить таким поведением. Если увидите, что ребенок спокоен и серьезен, используйте эту возможность, чтобы поговорить с ним о том, что его тревожит, беспокоит. Предложите ему нарисовать картинку, которую не увидит никто, кроме вас двоих. Если ребенок начинает баловаться в неподходящий момент и вам нужно прервать его фиглярство, твердо обратитесь к нему со словами: «Сейчас очень важно, чтобы ты побыл серьезным. Через 10 минут мы закончим работу, и ты сможешь баловаться, сколько пожелаешь».
Через 10 минут мы закончим работу, и ты сможешь баловаться, сколько пожелаешь».
Если ребенок плохо ест
Время еды должно быть периодом отдыха и удовольствия для всех. Тем не менее дети часто именно во время еды втягивают нас в конфликты и начинают самоутверждаться самым неподходящим способом.
Как предотвратить проблему
Вы являетесь для ребенка главным примером поведения за столом, поэтому должны есть вместе с ним и то же, что и он. Малышу следует предложить поесть, но ни в коем случае не заставлять есть насильно. Можно сказать: «Пожалуйста, скушай по маленькой порции каждого блюда, иначе это будет невежливо». Ребенок должен иметь право отказаться съесть «даже по маленькому кусочку». Если на него не давить, то рано или поздно он начнет есть нормально. Спокойный подход к еде способствует установлению здорового отношения к ней. Во время еды вовлекайте ребенка в неторопливый разговор. Почаще что-нибудь готовьте вместе.
Как справиться с проблемой
Если ребенок слишком быстро заглатывает пищу, накладывайте еду в тарелку понемногу, но обязательно скажите малышу, что приготовили еды достаточно и можете дать ему добавки. Объясните ребенку, что пищу нужно жевать медленно, чтобы организм мог взять из хорошо прожеванной пищи больше полезных веществ, это помогает человеку расти высоким, красивым и сильным.
Объясните ребенку, что пищу нужно жевать медленно, чтобы организм мог взять из хорошо прожеванной пищи больше полезных веществ, это помогает человеку расти высоким, красивым и сильным.
Если ребенок ест слишком вяло, накладывайте в тарелку пищу понемногу, постепенно добавляя ее. Объясните ему, что с маленькой порцией справиться гораздо легче. Не заостряйте на этой проблеме слишком много внимания, чтобы не усугубить ситуацию.
Если ребенок слишком привередлив в еде, для начала давайте ему новую еду очень маленькими порциями. Не заставляйте есть насильно. Постарайтесь задать за столом спокойный благожелательный тон, продемонстрируйте свое положительное отношение к конкретной еде.
Как нарисовать пшеницу: 3 способа
Пшеница — однолетнее зерновое растение, которое выращивают в большинстве стран мира. Из него делают муку, крупы, макароны и кондитерские изделия, даже пиво, и это далеко не полный список. А рисовать пшеницу совсем несложно. Ниже мы рассмотрим способы сделать это.
Как нарисовать пшеницу карандашом: материалы
Прежде чем приступить к рисованию пшеницы, подготовьте необходимые материалы. Это простые карандаши (лучше взять один твердый и один мягкий), лист бумаги, ластик и цветные карандаши (желтый, светло-оранжевый, зеленый, коричневый), если вы хотите раскрасить рисунок.
Первый путь
Для начала рассмотрим один из простейших способов поэтапного рисования пшеницы, который подходит для рисования с ребенком.
- На листе бумаги нарисуйте карандашом несколько слегка изогнутых вертикальных линий. Их количество зависит от того, сколько вы хотите нарисовать колоски.
- Нарисуйте девять кругов вокруг вершины одного из стеблей.
- Теперь нарисуйте круги из зерна, которые по форме выглядят как капли. Таким же образом нарисуйте волокна на оставшихся стеблях.
- На каждом колоске нарисуйте несколько усиков. Их должно быть немного, по 4-5 штук на каждом колоске. Снизу стеблей рисуем длинные и узкие листочки.

Теперь, когда вы знаете, как рисовать пшеницу простым способом, вы можете попробовать более сложный метод. В конце раскрасьте пшеницу оранжевым и желтым карандашами для колосков и зеленым карандашом для листьев.
Второй путь
Как нарисовать пшеницу другим простым способом? Для этого также потребуются лист бумаги, простой карандаш и ластик.
Рисуем тонкие, слегка наклонные линии. В верхней части каждой строчки изобразите колоски. Нарисуйте по несколько зерен с каждой стороны и по одной на самом верху. Зерна могут быть как в виде капель, так и в виде небольших овалов.
По обеим сторонам уха рисуем толстые полоски, а сверху рисуем несколько коротких линий. Дорисовываем пару листочков рядом со стеблями. Протираем ненужные линии — и рисунок готов. Можно нарисовать еще несколько колосков и раскрасить картину красками или карандашами, используя желтый и коричневый цвет.
Третий путь
Если вы хотите научиться рисовать пшеницу немного реалистичнее, то этот метод вам подойдет.
- Сплошным карандашом нарисуйте стебли со слегка изогнутыми линиями и вытянутыми овалами поверх них. На этом этапе будущие колоски напоминают камыши. При черчении сильно не давите на карандаш, линии должны быть немного видны.
- Далее внутри овалов рисуем крупинки, похожие на капли. Сначала рисуем одно зерно немного наклоненным, с другой стороны, второе чуть выше, третье снова чуть выше, напротив второго, и пока не заполняем нарисованный ранее овал.Не страшно, если вы выйдете немного за рамки контура.
- На концах зерен нарисуйте прямые линии ости.
- Теперь добавьте объема стеблям, проведя дополнительную линию рядом с существующей.
- Рисуем листья возле стеблей и стираем лишние линии. Чтобы изобразить сложенный кусок, нарисуйте вытянутый треугольник основанием вверх и под углом еще один такой же тонкий треугольник.
- Нарисуйте мягкий карандашный рисунок и добавьте небольшую тень у основания каждого зерна.Сверху и снизу слегка закрашиваем стебель и листья твердым карандашом.
 Можно использовать карандаш одинаковой твердости, нажимая на него с разной силой, чтобы получить разные оттенки. Также получившуюся картинку можно раскрасить.
Можно использовать карандаш одинаковой твердости, нажимая на него с разной силой, чтобы получить разные оттенки. Также получившуюся картинку можно раскрасить.
Таким образом можно нарисовать как одиночный колосок, так и целый сноп или даже поле. Кроме того, колоски можно немного модифицировать, разместив, например, зерна на немного большем расстоянии друг от друга, или нарисовав дополнительный ряд зерен сбоку.
рисунок из трех колосков ячменя на белом Фотография, картинки, изображения и сток-фотография без роялти. Изображение 61107514.
рисунок из трех колосков ячменя на белом Фотография, картинки, изображения и сток-фотография без роялти. Изображение 61107514. Рисунок трех колосков ячменя на белом.Колоски зерна пшеницы, изолированные на белом. Гравировка рисованной гравюры зерна злаков, ячменя и колоски для еды. Белый рисунок глубокой печати колосков и зерна пшеницы, изолированные на белом фоне. Рисованные колоски для печати на растениях ячменя.
Таблица размеров
| Размер изображения | Идеально подходит для |
| S | Интернет и блоги, социальные сети и мобильные приложения. |
| млн | Брошюры и каталоги, журналы и открытки. |
| л | Внутренние и наружные плакаты и печатные баннеры. |
| XL | Фоны, рекламные щиты и цифровые экраны. |
Используете это изображение на предмете перепродажи или шаблоне?
Распечатать Электронный Всесторонний
3508 x 4961 пикселей | 29.7 см x 42,0 см | 300 точек на дюйм | JPG
Масштабирование до любого размера • EPS
3508 x 4961 пикселей | 29,7 см x 42,0 см | 300 точек на дюйм | JPG
Скачать
Купить одно изображение
6 кредитов
Самая низкая цена
с планом подписки
- Попробовать 1 месяц на 2209 pyб
- Загрузите 10 фотографий или векторов.

- Нет дневного лимита загрузок, неиспользованные загрузки переносятся на следующий месяц
221 ру
за изображение любой размер
Цена денег
Ключевые слова
Похожие изображения
Нужна помощь? Свяжитесь со своим персональным менеджером по работе с клиентами
@ +7 499 938-68-54
Мы используем файлы cookie, чтобы вам было удобнее работать. Используя наш веб-сайт, вы соглашаетесь на использование файлов cookie, как описано в нашей Политике использования файлов cookie
Используя наш веб-сайт, вы соглашаетесь на использование файлов cookie, как описано в нашей Политике использования файлов cookie
колосков1.
- © Американское общество биологов растений, 2013. Все права защищены.
Abstract
Травы производят соцветия на структуре, называемой колоском, а разнообразие в количестве и расположении как ветвей, так и колосков способствует большому разнообразию архитектуры соцветий травы.У Brachypodium ( Brachypodium distachyon ) соцветие представляет собой неразветвленный колос с верхним колоском и ограниченным количеством боковых колосков. Колоски индетерминантные и дают различное количество цветков. Здесь мы приводим подробное описание этапов развития соцветий у Brachypodium. Чтобы получить представление о генетической регуляции развития соцветий Brachypodium, мы создали популяции мутантов с быстрыми нейтронами и проверили фенотипические мутанты. Среди идентифицированных мутантов мутант с большим количеством колосков1 ( mos1 ) имел повышенное количество пазушных меристем, продуцируемых из меристемы соцветия, по сравнению с диким типом. Эти пазушные меристемы развивались как ветви с образованием колосков более высокого порядка. Используя подход генов-кандидатов, было обнаружено, что mos1 имеет геномную перестройку, нарушающую экспрессию фактора ответа на этилен транскрипционного фактора APETALA2, связанного с генами идентичности меристемы колосков. mays ) и FRIZZY PANICLE ( FZP ) в рисе ( Oryza sativa ).Мы предполагаем, что MOS1 , вероятно, соответствует ортологу Brachypodium bd1 и FZP , и что функция этого гена в определении судьбы меристемы колосков сохраняется у отдаленно родственных видов трав. Однако MOS1 , по-видимому, также участвует во времени инициации терминального колоска. Таким образом, MOS1 может регулировать переход к развитию терминальных колосков у других близкородственных и важных с точки зрения сельского хозяйства видов, особенно пшеницы ( Triticum aestivum ).
Среди идентифицированных мутантов мутант с большим количеством колосков1 ( mos1 ) имел повышенное количество пазушных меристем, продуцируемых из меристемы соцветия, по сравнению с диким типом. Эти пазушные меристемы развивались как ветви с образованием колосков более высокого порядка. Используя подход генов-кандидатов, было обнаружено, что mos1 имеет геномную перестройку, нарушающую экспрессию фактора ответа на этилен транскрипционного фактора APETALA2, связанного с генами идентичности меристемы колосков. mays ) и FRIZZY PANICLE ( FZP ) в рисе ( Oryza sativa ).Мы предполагаем, что MOS1 , вероятно, соответствует ортологу Brachypodium bd1 и FZP , и что функция этого гена в определении судьбы меристемы колосков сохраняется у отдаленно родственных видов трав. Однако MOS1 , по-видимому, также участвует во времени инициации терминального колоска. Таким образом, MOS1 может регулировать переход к развитию терминальных колосков у других близкородственных и важных с точки зрения сельского хозяйства видов, особенно пшеницы ( Triticum aestivum ).
Побеги растений развиваются из апикальной меристемы побегов, которая включает центральную область плюрипотентных клеток и периферическую область, где клетки рекрутируются для формирования боковых органов. Апикальная меристема побега закладывается на ранних этапах эмбриогенеза и дает начало всем надземным частям растения. После репродуктивного перехода апикальная меристема побега превращается в меристему соцветия, которая может производить пазушные меристемы, образующие ветви и цветы. Производство, расположение и судьба меристем определяют архитектуру соцветий.
Архитектура соцветий растений очень разнообразна, и одно из наиболее ярких отражений этого разнообразия — в основных видах зерновых культур. У кукурузы ( Zea mays ) кисточка и колосовое соцветие неопределенные. Пазушные меристемы, образованные соцветием в кисточке, изначально представляют собой недетерминантные ветви. И кисточка, и колосовое соцветие, а также ветви кисточки образуют меристемы пары колосков, которые затем образуют определенные меристемы колосков, которые заканчиваются образованием двух цветков или соцветий (Bortiri and Hake, 2007; Kellogg, 2007).В соцветии или метелке риса ( Oryza sativa ) меристемы ветвей первоначально образуют вторичные меристемы ветвей. Как первичные, так и вторичные ветви образуют боковые меристемы колосков и терминальные меристемы колосков. Колосковая меристема дает единственную цветочную меристему и единственный цветочек (Bommert et al., 2005; Wang, Li, 2008). Для сравнения, соцветие или колос ячменя ( Hordeum vulgare ) и пшеницы ( Triticum aestivum ) не имеет ветвей (Bonnett, 1935, 1936).У ячменя меристема соцветия неопределенная и дает боковые меристемы колосков, образующие три соцветия. У пшеницы меристема соцветия детерминирована и дает ограниченное количество боковых меристем колосков и конечную меристему колосков, при этом каждая меристема колосков дает несколько цветков.
Гены, необходимые для развития соцветий, были идентифицированы путем анализа мутантов кукурузы и риса, и многие из этих генов контролируют инициацию и судьбу меристемы (Bommert et al., 2005; Sreenivasulu and Schnurbusch, 2012). Одна группа мутантов не может давать ветки и колоски. бесплодный стебель1 ( ba1 ) у кукурузы приводит к радикальному снижению продукции пазушных меристем и образует неразветвленное соцветие без колосков (Ritter et al., 2002; Gallavotti et al., 2004). Мутация в LAX PANICLE1 ( LAX1 ), ортологе риса ba1 , также приводит к меньшему количеству колосков (Komatsu et al., 2001, 2003b).Ген риса MONOCULM1 ( MOC1 ) (также известный как SMALL PANICLE ) необходим для инициации подмышечных меристем на протяжении вегетативного и репродуктивного развития. Мутанты moc1 лишены побегов и имеют мало ветвей и колосков соцветий (Li et al., 2003; Oikawa, Kyozuka, 2009). бесплодное соцветие2 ( bif2 ) кукурузы не дает ветвей или колосков в соцветии и требуется для поддержания всех типов пазушных меристем (McSteen and Hake, 2001). ba1 , LAX1 и MOC1 кодируют факторы транскрипции, тогда как bif2 кодирует белок протеинкиназы Ser / Thr, участвующий в передаче сигналов ауксина (Komatsu et al., 2003b; Li et al., 2003; Gallavotti et al. ., 2004; McSteen et al., 2007).
Другая группа мутантов кукурузы и риса усиливает ветвление соцветий. Мутанты кукурузы ramosa1 ( ra1 ), ra2 и ra3 меняют меристемы из пары колосков на меристемы ветвей, что приводит к увеличению количества ветвей в кисточке и развитию ветвей в колосе (Vollbrecht et al., 2005; Bortiri et al., 2006; Сато-Нагасава и др., 2006). Аналогичным образом, мутанты в семействе ортологичного фактора этиленового ответа (ERF) генов фактора транскрипции APETALA2 (AP2) разветвленного без шелка1 ( bd1 ) в кукурузе и FRIZZY PANICLE ( FZP ) в соцветиях риса приводят к сильному разветвлению. к превращению меристем колосков в меристемы ветвей (Colombo et al., 1998; Chuck et al., 2002; Komatsu et al., 2003a; Zhu et al., 2003; Yi et al., 2005).
Хотя многие гены архитектуры соцветий были идентифицированы у видов трав с разветвленными соцветиями, гораздо меньше известно о факторах, регулирующих развитие неразветвленных соцветий (Malcomber et al., 2006). В неразветвленном соцветии, известном как колос, пазушные меристемы, образованные меристемой соцветия, развиваются непосредственно в колоски. Как и у пшеницы и ячменя, соцветие Brachypodium ( Brachypodium distachyon ) представляет собой колос (Draper et al., 2001; Vogel et al., 2006). Чтобы понять регуляцию развития колючек, мы провели исследование стадий развития колосков Brachypodium. Здесь мы представляем подробный анализ онтогенеза соцветия, поскольку он связан с развитием меристемы у Brachypodium дикого типа (номер доступа Bd21). Кроме того, чтобы начать изучение генетической регуляции архитектуры соцветий Brachypodium, мы создали условия для быстрых нейтронов Brachypodium и химического мутагенеза. Популяции мутантов быстрых нейтронов имели высокую частоту мутантов, и из этих популяций мы идентифицировали ряд мутантов.Три мутанта по соцветию, безостый1 ( awl1 ), без соцветия1 ( nif1 ) и бесхвостый1 ( til1 ), имели дефектное развитие колосков и / или цветков и еще у одного мутанта . ( mos1 ), колоски переделали в ветки. Мы показываем, что дефект соцветия mos1 является результатом изменений судьбы меристемы колосков во время развития соцветий. Наконец, мы показываем, что mos1 , вероятно, соответствует мутации в гене Brachypodium, наиболее тесно связанной с кукурузой bd1 и рисом FZP .Таким образом, bd1 , FZP и MOS1 обладают консервативной функцией, чтобы способствовать детерминированной судьбе меристемы колосков в травах. Однако, MOS1 в Brachypodium, по-видимому, участвует в инициации терминального колоска и, таким образом, может контролировать количество колосков, образующихся в неразветвленном детерминированном колоске.
РЕЗУЛЬТАТЫ
Развитие соцветий Brachypodium
Чтобы заложить основу для описания развития соцветий Brachypodium, мы провели детальный анализ онтогенеза соцветий дикого типа.После прорастания первый настоящий лист был виден через 6 дней после посева, а шесть-семь дополнительных листьев сформировались до появления меристемы соцветия, примерно через 22 дня после посева (рис. 1A). Подмышечные меристемы у основания растения удлиненные, образуя несколько базальных стеблей, похожих на основной стебель (стебель; рис. 1А). Степень образования побегов варьировалась в зависимости от условий произрастания. При более низкой освещенности образовалось больше побегов, и в этих условиях переход к цветению задерживался.
Рисунок 1.Развитие соцветий Brachypodium. А. Целые растения через 6, 10, 14, 18, 22 и 26 дней после посева (дас). От B до E: продольные срезы верхушки побега растений через 6 дней (B), 10 дней (C), 14 дней (D), 18 дней (E). am, подмышечная меристема; fm — цветочная меристема; ls, боковой колоск; sam — апикальная меристема побега; см — меристема колосков; ц, концевой колоск. Полосы = 2 см (A) и 100 мкм (B – E). [Цветную версию рисунка см. В статье в Интернете.]
Соцветие Brachypodium состояло из двух или трех боковых колосков и концевого колоска (рис.2А). Каждый колосок состоял из двух базальных чешуек и в среднем из 11 цветков, расположенных двояковыпуклой филлотаксией вдоль центрального стержня (рис. 2В; табл. I). Как и при выращивании культиватора, количество колосков и цветков, а также завязка семян зависели от условий роста. При менее чем оптимальных условиях роста плодовитость была низкой, и в этом случае колос давал больше цветков. Цветки состояли из наружной остистой леммы, полупрозрачной палеи, двух лодикул, трех тычинок и центрального пестика (рис.2, В и Г). Две боковые тычинки созрели и дали пыльцу, тогда как абаксиальные тычинки остались рудиментарными и стерильными.
Рисунок 2.Развитие колосков и цветков Brachypodium. А, соцветие дикого типа с концевыми и боковыми колосками. B, Терминальный колоск. C, Флоре показывает палею (слева) и лемму с дистальной остью (справа). D, СЭМ репродуктивных органов цветков. Палеа была удалена, чтобы обнажить два развитых пыльника и пестик. E, Ранняя голая стадия развития соцветия с зарождающейся цветочной меристемой терминальных и боковых колосков.F, стадия инициации ости, когда рост цветковых органов в терминальном колоске более развит у базальных цветков по сравнению с апикальными цветками. G, закрытая стадия терминального колоска с наружной чешуей терминального колоска, окружающего развивающиеся соцветия. H, гибридизация in situ экспрессии BdKN1 в колоске. I. Филограмма максимального правдоподобия белков KNOX класса I Brachypodium, риса и кукурузы. Значения начальной загрузки максимального правдоподобия указаны на ветвях. а, Awn; ан, пыльник; ф, цветочек; fm — цветочная меристема; gl, чешуйка; lgl, нижняя чешуя; ls, боковой колоск; l, лемма; п — пестик; ts, терминальный колоск; цм — терминальная меристема колосков; угл, верхняя чешуя.Штанги = 100 мкм. [Цветную версию этого рисунка см. В статье в Интернете.]
Таблица I. Число колосков и цветочков в колосе BrachypodiumЧтобы определить время перехода от вегетативного к репродуктивному периоду, мы подготовили продольные срезы верхушек растений и исследовали морфологию побегов апекс от 6-, 10-, 14- и 18-дневных растений, выращенных в условиях длинного дня. У 6-дневных растений апикальная меристема побега представляла собой короткий купол клеток, по бокам которого развивались зачатки листьев (рис.1Б). У 10-дневных растений апикальная меристема была более удлиненной, чем у 6-дневных растений, и начинался концевой колоск (рис. 1С). Таким образом, переход к репродуктивной судьбе происходил у растений возрастом от 7 до 9 дней. Цветочные меристемы терминального колоска проявлялись на верхушках 10-, 14- и 18-дневных растений на продольных срезах, тогда как боковые колоски с явными цветочными меристемами были заметны только на более поздней стадии у 18-дневных растений ( Рис.1, D и E). Следовательно, развитие терминального колоска предшествует развитию боковых колосков.
Чтобы охарактеризовать более поздние стадии развития соцветий Brachypodium, мы вскрыли верхушки 17–22-дневных растений и исследовали развитие ранних соцветий с помощью сканирующей электронной микроскопии (СЭМ). Меристема соцветия образовывала боковые колоски в виде дистихической филлотаксии, тогда как концевой колоск был ориентирован под углом 90 ° к плоскости боковых колосков (рис. 2E). В соответствии с результатами продольных срезов, анализ SEM показал, что развитие терминального колоска было более развитым, чем развитие боковых колосков (рис.2, E – G). Таким образом, развитие соцветия легче всего проследить путем осмотра терминального колоска. На самой ранней исследованной стадии развития соцветий терминальные колоски инициировали несколько цветочных меристем (рис. 2E). Инициирование самого внешнего цветочного органа, леммы, было очевидным как гребень клеток в основании цветочных меристем, тогда как другие цветочные органы не были очевидны. Две чешуйки, связанные с терминальным колоском, также образовались, причем верхняя чешуйка более заметна, а нижняя чешуя представлена только в виде небольшого гребня ткани.Мы назвали это «голой» стадией развития соцветия, так как цветочные меристемы терминального колоска были полностью обнажены. Следующая стадия развития наступила, когда зародились несколько цветков терминального колоска и характеризовалась ростом чешуек и нижней части базальных цветков (рис. 2F). Развивающаяся лемма вытянулась и дифференциальный рост дистальной средней области леммы привел к образованию ости. Мы классифицировали это как стадию «инициации ости». Впоследствии верхняя чешуйка и самый базальный цветочек удлинились, чтобы покрыть терминальный колоск, и это произошло до удлинения латеральных чешуек колосков (рис.2G; Дополнительный рис. S1). Мы назвали это стадией «закрытого концевого колоска».
Как в концевых, так и в боковых колосках цветочные меристемы развивались акропетально (рис. 2F). Чтобы подтвердить прогрессирование развития меристемы цветков в терминальном колоске и установить информативный молекулярный маркер для развития меристемы соцветия, мы использовали последовательность всего генома для идентификации генов Brachypodium, кодирующих факторы транскрипции KNOX I класса. Гены -подобных гомеобоксов (KNOX) класса I необходимы для функции меристемы побегов, экспрессируются в меристемах побегов и подавляются в боковых органах (Jackson et al., 1994; Long et al., 1996; Sentoku et al., 1999). Brachypodium имеет семь генов KNOX класса I (Vogel et al., 2010). Филогенетический анализ с максимальной вероятностью белков KNOX класса I Brachypodium, кукурузы и риса показал, что Brachypodium кодирует один ортолог в каждой кладе гена KNOX класса I, за исключением клады knox3, где ген Brachypodium отсутствует, и клады knox10, где есть два гена, Bradi1g12677 и Bradi1g12690 , которые расположены рядом друг с другом на хромосоме 1 (рис.2I). По сравнению с другими генами KNOX класса I, Bradi1g12677 усечен и может быть псевдогеном.
Белок, кодируемый Bradi1g10047, был наиболее тесно связан с KNOTTED1 (KN1) кукурузы и риса HOMEOBOX1 (OSh2; рис. 2I). Таким образом, Bradi1g10047 получил обозначение BdKN1 . Мы использовали гибридизацию in situ для изучения паттерна экспрессии BdKN1 в развивающемся терминальном колоске Brachypodium дикого типа и обнаружили, что он высоко экспрессируется в меристемах колосков и цветков.(Рис. 2H). Подобно kn1 , экспрессия BdKN1 не была обнаружена во внешнем слое клеток L1 всех меристем побегов. BdKN1 подавляется в небольшой области, смежной с терминальной меристемой колосков, которая соответствует участку инициирующего органа. От вершины к основанию колоска цветочные меристемы становились все больше и BdKN1 подавлялся в развивающихся органах, ассоциированных с этими меристемами. Таким образом, паттерн экспрессии BdKN1 совпадает с паттерном экспрессии близкородственных генов у других растений и маркирует активные меристемы у Brachypodium.Характер экспрессии также подтверждает акропетальное созревание цветков колосков.
Мутагенез Brachypodium
Чтобы идентифицировать гены, регулирующие развитие соцветий у Brachypodium, мы создали мутагенизированные популяции, которые были проверены на наличие мутантов по развитию соцветий. Мы использовали два подхода к мутагенезу, химический мутагенез этилметансульфоната (EMS) и облучение быстрыми нейтронами, которые оказались успешными в создании мутантов у других видов. Мутанты были получены с использованием обоих подходов (дополнительный рис.S2). Популяции, подвергшиеся мутагенезу быстрыми нейтронами, давали более частые мутанты и поэтому стали предметом последующего скрининга.
Два испытания мутагенеза на быстрых нейтронах продемонстрировали, что воздействия от 20 до 30 Грея (Гр) было достаточно для получения растений, которые дают жизнеспособные семена (дополнительный рис. S2, A и B). Используя эти две независимые популяции мутантов с быстрыми нейтронами, мы провели скрининг потомства M2 из 872 линий M1 на наличие видимых фенотипов побегов. Мутанты были разделены на три основные категории: альбиносы, бледные и с дефектами развития (дополнительные рис.S2, C, D и S3). В двух популяциях мутантов было 5% линий, разделяющих мутантов-альбиносов, и 4% линий, разделяющих бледных мутантов (дополнительный рисунок S3A). Класс мутантов с дефектным развитием охватывает фенотипы, нарушающие развитие побегов растений, и эти мутанты были идентифицированы в 2% и 4% линий в двух популяциях мутантов. Мутанты развития включали растения, которые были чрезвычайно карликовыми мутантами, мутант с морщинистым листом и мутант с нисходящим ростом (дополнительный рис. S3, B – D).Частота мутантных фенотипов, особенно высокая частота мутантов-альбиносов в линиях быстрых нейтронов, указывает на то, что доза от 20 до 30 Гр эффективна для мутагенеза Brachypodium.
Три мутанта: awl1 , nif1 и til1 нарушили развитие соцветий. awl1 был подобен дикому типу во время вегетативного развития; однако при репродуктивном развитии цветковая перепонка не образовывала ости, а развитие пестика было аномальным (рис.3, А – Д). nif1 также был подобен дикому типу во время вегетативного развития; однако при переходе к развитию соцветий nif1 часто не образовывал колосков и вместо этого давал голый стебель (рис. 3, F и H). Мутант til1 был нарушен как в вегетативном развитии, так и в развитии соцветий. У дикого типа побеги возникают в пазухах листьев у основания растения, тогда как у мутанта til1 побеги отсутствуют (рис. 3G).В соцветии til1 конечный колоск и боковые колоски имели дефектные и уменьшенное количество цветков (рис. 3, G и I). Четвертый мутант, mos1 , также влиял на развитие соцветий и давал больше колосков, чем у дикого типа. Этот мутант был выбран для дальнейшего анализа.
Рисунок 3.Мутанты по соцветию. A, awl1 цветковое растение. B и C, Spike дикого типа (B) и awl1 (C). D и E, Репродуктивные органы дикого типа (D) и awl1 (E).Пыльники отмечены стрелками. F и G, nif1 (F) и til1 (G) цветущее растение. H и I, шипы в nif1 (H) и til1 (I). [См. Онлайн-статью для цветной версии этого рисунка.]
mos1 превращает колоски в ветвиМутант mos1 был фертильным и выделился как рецессивный мутант. Во время вегетативного роста mos1 существенно не отличался от дикого типа и отличался от дикого типа только по соцветию (рис.4, А и Б). Однако во время репродуктивного роста соцветие давало примерно в два раза больше боковых колосков по сравнению с диким типом (рис. 4С; таблица II). Боковые колоски mos1 не были покрыты чешуей и поэтому считались ветвями. Каждая ветвь имела в среднем от 2,2 до 3,9 колосков, включая конечный колоск и первичные боковые колоски (Таблица II). Ветви давали несколько порядков колосков, так что одна ветвь давала первичные боковые колоски, которые, в свою очередь, давали вторичные боковые колоски (рис.4, Г и Д). Ветвистый верхний колосок также давал в среднем три вторичных колоска, и у них было в среднем 7,6 цветков, что меньше, чем количество цветков в колоске дикого типа (Таблицы I и II). Пороков органов у цветков не было. Колоски у основания ветвей, как правило, были небольшими с рудиментарными цветочками с уменьшенным количеством органов (Рис. 4C; Таблица II). Таким образом, mos1 приводит к разветвлению колоса Brachypodium.
Рис. 4. Мутантmos1 .A и B, Цветущее растение дикого типа (A) и mos1 (B). C, mos1 шип. Цифрами обозначены верхний колоск (1) и ветви (2–6). Д, Мос1 филиал. Стрелками обозначены первичные боковые колоски. E, mos1 первичный боковой колоск. Стрелками обозначены вторичные боковые колоски. rs, рудиментарный колоск. [См. Онлайн-статью для цветной версии этого рисунка.]
Таблица II. Количество ветвей, первичных колосков и цветков в соцветии mos1mos1 развитие соцветий было дополнительно охарактеризовано с помощью SEM.На голой стадии развития боковые ветви казались похожими на колоски дикого типа, и каждая ветвь инициировала подмышечные меристемы, подчиненные развивающемуся органу (рис. 5, A и B). Когда ветви переходили в стадию зарождения ости, пазушные меристемы ветвей образовывали боковые колоски (первичные боковые колоски; рис. 5C). На каждой ветке образовалось от двух до четырех первичных боковых колосков (табл. II). Конечный колоск ветви можно было различить, так как он был покрыт чешуей и был более развит в развитии, чем боковые колоски.
Рисунок 5. Развитие соцветияmos1 . A и B, обнаженная сцена. C, Ветвь на стадии инициации ости. D, соцветие, заключенное в конечную стадию колоска, с концевым колоском (1) и пятью ветвями. а, Awn; am, подмышечная меристема; б, ветвь; gl, чешуйка; l, лемма; плс, первичный боковой колоск; ц, концевой колоск. Штанги = 100 мкм.
Как и в соцветии дикого типа, у соцветия mos1 развитие концевого колоска было более развитым, чем у боковых ветвей (рис.5D). Кроме того, были выражены различия в созревании ветвей вдоль соцветия. На стадии развития соцветий с закрытым концевым колоском четыре-шесть базальных ветвей находились на стадии зарождения ости, а две-три верхушечные ветви находились на стадии голого развития. MOS1 поэтому, по-видимому, участвует в производстве латеральных колосков, способствуя переходу меристемы соцветия в терминальный колоск и способствуя переходу пазушных меристем, продуцируемых соцветием, в судьбу колосков.
mos1 несет хромосомную перестройку перед геном фактора транскрипции ERF, относящимся к bd1Ортологичные гены ERF bd1 кукурузы и FZP в рисе необходимы для сохранения идентичности меристемы колосков. , 2002; Komatsu et al., 2003a; Zhu et al., 2003). Мутации в обоих генах приводят к превращению детерминированных меристем колосков в меристемы неопределенных ветвей, напоминающих mos1 .Учитывая это сходство, мы проверили, может ли mos1 быть нарушен в гене, родственном Brachypodium bd1 . BD1 и FZP являются факторами транскрипции ERF, и существует 142 фактора транскрипции AP2 / ERF, кодируемых в геноме Brachypodium (Vogel et al., 2010). Используя аминокислотные последовательности BD1 и FZP для поиска аннотированных последовательностей белка Brachypodium, мы обнаружили только один близкородственный белок, кодируемый Bradi1g18580, который имеет общую аминокислотную идентичность в домене ERF и выходит за пределы этого домена (рис.6А; Vogel et al., 2010). Мы также идентифицировали Sb02g042400 из сорго ( Sorghum bicolor ) и предполагаемый белок в последовательности генома ячменя как наиболее близкий родственник BD1 и FZP (Paterson et al., 2009; Mayer et al., 2012). Белок Bradi1g18580 имел 59%, 54%, 57% и 58% идентичности с BD1, FZP, Sb02g042400 и белком ячменя соответственно (фиг. 6A). Филогенетический анализ максимального правдоподобия этих белков, связанных с BD1, соответствовал взаимоотношениям между этими видами и показал, что белки Brachypodium и ячменя наиболее тесно связаны между собой (дополнительный рис.S4A; Келлогг, 2001; Vogel et al., 2006). Чтобы проверить, был ли этот кандидат разрушен в mos1 , Bradi1g18580 был амплифицирован с помощью ПЦР от дикого типа и mos1 . Секвенирование продуктов ПЦР не выявило мутаций в кодирующей области Bradi1g18580. Однако ПЦР-амплификация 5′-области Bradi1g18580 выявила разницу в размере продукта между продуктом дикого типа и mos1 . Секвенирование этих продуктов амплификации выявило делецию 10 п.н., простирающуюся от -170 до -180 п.н. выше стартового кодона ATG Bradi1g18580, а также уникальную вставку последовательности 991 п.н. на -170 п.н. выше стартового кодона ATG в mos1. относительно дикого типа (рис.6, Б и В). Интересно, что вставка была идентична последовательности на хромосоме 3, предполагая, что мутация возникла в результате хромосомной перестройки, когда сегмент хромосомы 3 вставлен в хромосому 1. Анализ последовательности показал, что вставка хромосомы 3 не имела признаков мобильного элемента и не имела иметь любые открытые рамки считывания, указывающие на последовательность, кодирующую белок. Чтобы определить, может ли эта вставка быть ответственной за фенотип mos1 , мы проверили косегрегацию между фенотипом mos1 и вставкой.В линиях, разделяющих mos1 , все 108 исследованных мутантов были гомозиготными по инсерционному аллелю, и все 36 исследованных фенотипических растений дикого типа либо не несли инсерционный аллель, либо были гетерозиготными по инсерционному аллелю и инсерционному аллелю. Основываясь на опубликованных частотах рекомбинации у Brachypodium, вставки выше Bradi1g18580 и mos1 , по оценкам, связаны менее чем на 100 kb (Huo et al., 2011).
Рисунок 6.MOS1 соответствует мутации в ортологе Brachypodium bd1 и FZP .A. Выравнивание аминокислотных последовательностей предсказанных белков из Brachypodium MOS1 (белок, кодируемый Bradi1g18580), кукурузы BD1, FZP риса и ортологов BD1 сорго и ячменя. Цветовое затенение указывает процентную идентичность. Домен ERF подчеркнут. B, продукты амплификации ПЦР дикого типа и mos1 , амплифицированные с праймерами, показанными на C. Праймеры F5 и R5 обнаруживают полиморфизм вставки в mos1 . C, схематическое изображение Bradi1g18580 в mos1 с единственной открытой рамкой считывания (зеленый прямоугольник) и вставкой хромосомы 3 (красная линия).Цифры — это нуклеотиды, при этом цифра 1 — начало открытой рамки считывания. Стрелки указывают расположение праймеров. Нуклеотидная последовательность показывает область, несущую перестройку. Нуклеотиды красного цвета — это концы вставки хромосомы 3. Нуклеотиды, выделенные серым цветом, удалены в mos1 . Желтые прямоугольники выделяют нуклеотиды, которые сохраняются в Brachypodium, ячмене, кукурузе, рисе и сорго. D, количественная ОТ-ПЦР, показывающая уровни транскриптов MOS1 по сравнению с ACTIN2 у дикого типа и мутанта mos1 .Полоски = среднее значение.
Вставка в Bradi1g18580 нарушает высококонсервативную последовательность в Brachypodium, ячмене, рисе, кукурузе и сорго, включая сайт начала транскрипции ТАТА-бокса (рис. 6C; дополнительный рис. S4B). Чтобы определить, изменяет ли вставка уровни транскрипции Bradi1g18580, мы выполнили количественную обратную транскрипцию (RT) -PCR и обнаружили, что уровни транскрипта Bradi1g18580 были снижены в mos1 по сравнению с диким типом (фиг. 6D). Умеренное снижение уровней транскрипта MOS1 у мутанта указывает на то, что mos1 является слабым аллелем, который согласуется с мягким фенотипом mos1 по сравнению с аллелями bd1 и fpz .Взятые вместе, генетические, фенотипические и транскрипционные данные предполагают, что MOS1 соответствует Bradi1g18580.
ОБСУЖДЕНИЕ
Мутанты быстрых нейтронов в Brachypodium
Brachypodium представляет собой полезную модельную систему для исследования генетической регуляции развития колосков, поскольку это быстрорастущий и легко культивируемый вид с относительно небольшим геномом, который недавно был секвенирован (Draper et al. al., 2001; Vogel et al., 2010; Brkljacic et al., 2011). Мы создали условия для мутагенеза быстрых нейтронов и EMS.Скрининг популяций мутантов быстрых нейтронов выявил ряд фенотипических мутантов. Некоторые из мутантов онтогенеза имели фенотипы, сходные с мутантами, о которых сообщалось у других видов растений. Например, мутант с растущим вниз побегом был подобен агравитрофным мутантам lazy в кукурузе и рисе и serpentina в ячмене (Van Overbeek, 1936; Jones and Adair, 1938; Sheridan, 1988; Türkan and Suge, 1991). ).
Выявлен ряд мутантов с дефектами развития соцветий.Мутант awl1 не имел ости и имел дефектное развитие пестика. Оости присутствуют в других видах трав, таких как рис, ячмень и пшеница, и, вероятно, выполняют функцию защиты от кормления животных и способствуют распространению семян, хотя ости также могут играть физиологическую роль в развитии семян (Grundbacher, 1963). Хотя еще предстоит определить, контролирует ли один-единственный ген развитие ости и пестика, мутант awl1 обеспечит понимание обоих процессов в развитии соцветий.
Мутанты nif1 и til1 имели сходство с другими описанными мутантами. Мутант nif1 , у которого отсутствовали колоски, напоминал мутанты транспорта ауксина , сформированные1 , пиноидные и моноплодные у арабидопсиса ( Arabidopsis thaliana ) и bif2 у кукурузы (Okada et al., 1991; al., 1995; Przemeck et al., 1996; McSteen et al., 2007). PINFORMED1 кодирует переносчик оттока ауксина, который экспрессируется на высоких уровнях в локализованных областях на периферии меристемы и необходим для образования локальных максимумов ауксина, связанных с инициацией органа (Reinhardt et al., 2003). Аналогичным образом, мутанты в bif2 , который кодирует протеинкиназу Ser / Thr, ортологичную PINOID , имеют меньше ветвей соцветий, колосков и органов цветков и не могут установить локализованные максимумы ауксина в периферических областях меристемы (McSteen et al. др., 2007; Скирпан и др., 2009). Потенциально nif1 также является дефектным в транспорте ауксина. Фенотип til1 был подобен фенотипу moc1 у риса и мутантам в ортологах ba1 кукурузы и LAX1 риса. MOC1 кодирует ген фактора транскрипции семейства GRAS, тогда как ba1 и LAX1 кодируют ортологичные основные факторы транскрипции спираль-петля-спираль (Komatsu et al., 2003b; Li et al., 2003; Gallavotti et al., 2004 ). Ортологи Brachypodium этих генов, следовательно, представляют собой кандидатов на TIL1 .
Сравнение развития соцветий у брахиподиума с мелкозерновыми злаками Пшеница и ячмень
Брахиподиум является членом подсемейства Pooideae семейства Poaceae и тесно связан с двумя основными видами мелкозерновых злаков — пшеницей и ячменем (Hsiao et al. al., 1994; Vogel et al., 2006). Основываясь на нашем анализе Brachypodium дикого типа, информативно сравнить это с развитием соцветий у пшеницы и ячменя, поскольку генетическая регуляция развития соцветий, вероятно, будет в высокой степени консервативной у этих родственных видов. Соцветие всех трех видов — колосовидное. Колосья ячменя характеризуется неопределенным соцветием и детерминированными колосками (Bonnett, 1935; Kirby, Appleyard, 1981; Babb, Muehlbauer, 2003). Пазушные меристемы вдоль главной оси соцветия дают три колоска, и каждый из этих колосков дает единственный цветочек.У пшеницы соцветие детерминантное, и меристема соцветия продолжает продуцировать пазушные меристемы до тех пор, пока не сформируется терминальный колоск (Bonnett, 1936; Kirby and Appleyard, 1981). Каждая пазушная меристема на соцветии пшеницы развивается в один боковой колоск. И верхние, и боковые колоски дают несколько цветков. Колос соцветия Brachypodium похож на колос пшеницы, при этом соцветие однозначное, а колоски неопределенные.Более того, соцветия в колоске созревают акропетально как у Brachypodium, так и у пшеницы. Однако у пшеницы наблюдается градация созревания колосков по оси соцветия. Колоски развиваются акропетально и базипетально из средней части соцветия, причем последний созревает последним (Bonnett, 1935, 1936; Kirby, Appleyard, 1981). Напротив, развитие терминального колоска у Brachypodium более развито по сравнению с латеральными колосками. Сложнее сделать выводы об относительном графике созревания нескольких боковых колосков у Brachypodium дикого типа.Однако у mos1 развитие базальных боковых ветвей было более развитым, чем развитие апикальных боковых ветвей, это указывает на то, что развитие вдоль главной оси соцветия у Brachypodium является акропетальным. Таким образом, есть поразительное сходство между развитием соцветий у Brachypodium и пшеницы, но тонкие различия могут отражать различную регуляцию созревания меристемы, а не зарождение.
mos1 преобразовывает неразветвленный шип в разветвленный шипМы создали описание развития шипа дикого типа Brachypodium и использовали его для определения роли MOS1 в архитектуре шипа.Мутация в MOS1 приводит к увеличению числа пазушных меристем на главной оси соцветия, которые развиваются скорее в виде ветвей, чем колосков. Повышенное количество подмышечных меристем может быть результатом задержки в формировании терминального колоска. MOS1 , следовательно, может либо способствовать продукции терминального колоска и определять судьбу меристемы, либо репрессировать неопределенную судьбу меристемы соцветия. Боковые пазушные меристемы в соцветии mos1 не покрыты чешуей и поэтому могут считаться ветвями, а не колосками.Каждая ветка напоминает главное соцветие дикого типа, образуя несколько боковых колосков и верхний колоск. Таким образом, идентичность меристемы колосков преобразуется в идентичность меристемы ветвления и аналогична фенотипам мутантов bd1 и fzp кукурузы и риса.
Мутантные аллели bd1 и fzp различаются по степени тяжести фенотипа. У тяжелых мутантов bd1 , мутации которых предсказывают усечение белка BD1, колоски кисточки являются неопределенными и образуют серию боковых колосков, а колоски ушей превращаются в неопределенные ветви (Chuck et al., 2002). Слабый аллель со сниженным уровнем транскрипта из-за вставки транспозона в 5′-лидерную последовательность bd1 приводит к образованию меньшего количества ветвей в ухе по сравнению с тяжелыми аллелями и является частично фертильным (Chuck et al., 2002). У тяжелых мутантов fzp все колоски на первичных ветвях метелки превращаются во вторичные ветви, тогда как у растений со слабым аллелем fzp колоски и иногда плодовитые соцветия образуются на концах первичных ветвей (Komatsu et al. ., 2001, 2003а; Чжу и др., 2003; Йи и др., 2005). Слабый аллель fzp имеет аминокислотную замену в ERF-домене FZP. Тяжелые аллели fzp имеют мутации, которые изменяют аминокислоты в домене ERF или, как предполагается, продуцируют C-концевые усеченные белки. Кроме того, два тяжелых мутанта fzp несут транспозонные вставки более чем на 1 т.п.н. выше кодирующей области, что, вероятно, нарушает экспрессию FZP через отдаленную цис-регуляторную область (Komatsu et al., 2003a; Чжу и др., 2003). Таким образом, помимо функции белка, регуляция транскрипции bd1 и FZP важна для образования колосков и для определения степени ветвления соцветий кукурузы и риса. Как и в случае слабых аллелей bd1 и fzp , мутанты mos1 являются фертильными. mos1 имеет хромосомную перестройку в промоторной области гена, и уровни транскриптов снижены по сравнению с диким типом. Поэтому мы предполагаем, что mos1 может быть слабым аллелем, фенотип которого является результатом более низких уровней накопления транскриптов или задержки начала экспрессии.
Неразветвленный колос Brachypodium отличается от разветвленного соцветия кукурузы и риса, и один ген, который может способствовать этому различию, — это bd1 / FZP / MOS1 . Мы показываем, что MOS1 , вероятно, соответствует мутации, которая приводит к снижению уровней транскрипта в ортологе Brachypodium bd1 и FZP . Дальнейшее снижение экспрессии MOS1 посредством выделения дополнительных мутантных аллелей или посредством использования сайленсинга генов подтвердит этот вывод, а также покажет степень, в которой MOS1 требуется для детерминированности меристемы.В частности, фенотип нулевого мутанта mos1 будет определять, зависит ли спецификация концевого колоска от экспрессии MOS1 . Изменения в уровне или характере экспрессии этого гена могут определять, является ли соцветие травы разветвленным или нет. Хотя bd1 , FZP и MOS , по-видимому, играют консервативную роль в судьбе меристемы колосков, мы показали, что MOS1 в Brachypodium также играет роль в детерминации меристемы соцветия.Одна из возможностей состоит в том, что MOS1 регулирует время инициации терминального колоска. У пшеницы дифференциация верхних колосков определяет количество образовавшихся колосков (Bonnett, 1935, 1936). MOS1 , таким образом, является хорошим геном-мишенью для генетических манипуляций с целью увеличения количества зерен и урожайности пшеницы.
МАТЕРИАЛЫ И МЕТОДЫ
Растительный материал и условия роста
Brachypodium ( Brachypodium distachyon ) Bd21, используемый в качестве дикого типа, был десятым поколением односемянного происхождения и выращивался либо в камере для выращивания, либо в теплице на 22 ° C при 18-часовом освещении.Семена высевали на влажную фильтровальную бумагу, помещали при 4 ° C в темноте на 7 дней и проращивали либо при комнатной температуре, либо в камере для выращивания в течение 4 дней перед пересадкой в почву, состоящую из 50% компоста и 50% торфа и песка. смесь с добавлением удобрений с медленным высвобождением. Подробные фенотипические исследования дикого типа и мутантов были проведены на растениях, выращенных в камере для выращивания.
Мутагенез на быстрых нейтронах семян Brachypodium был проведен в Исследовательском институте атомной энергии в Будапеште, Венгрия.Для определения дозы, необходимой для мутагенеза быстрых нейтронов, семена Brachypodium первоначально подвергались воздействию 20, 30, 40, 50 и 60 Гр. Семена M1 проверяли на жизнеспособность и плодовитость проростков. Семена, подвергнутые дозе 50 и 60 Гр, все прорастали, но не давали никаких постэмбриональных органов. Воздействие 40 Гр привело к прорастанию 25% семян M1 и росту проростков, а 22% растений были фертильными. Воздействие 30 Гр обеспечило прорастание 65% семян M1, и 60% растений были фертильными. На основе этого первоначального теста был проведен второй мутагенез, и семена подверглись воздействию 20, 25, 30 и 35 Гр.Семена, подвергнутые воздействию 35 Гр, имели низкую жизнеспособность проростков, и все созревшие растения были стерильными. Воздействие 20, 25 и 30 Гр обеспечило от 30% до 50% жизнеспособности проростков. Только при 20 Гр наиболее жизнеспособные сеянцы оплодотворяли. Были собраны семена от отдельных плодородных растений M1. Мы проверили потомство M2 от 203 линий M1, подвергшихся воздействию 30 Гр из первого мутагенеза, и 669 линий M1, подвергшихся воздействию 20 и 30 Гр из второго мутагенеза, на предмет видимых фенотипов побегов. Для скрининга мутантных фенотипов примерно 40 семян M2 от каждого растения M1 проращивали и высаживали в почву для визуального исследования.
EMS-мутагенез осуществляли путем сначала пропитывания очищенных от шелухи семян, а затем обработки EMS. Очищенные семена пропитывали, помещая на влажную фильтровальную бумагу на 48 часов при 4 ° C. Семена собирали и обрабатывали 0,2%, 0,4% или 0,6% (об. / Об.) EMS в 40 мл воды в течение 8 часов при комнатной температуре. В качестве контроля некоторые семена обрабатывали без EMS. После химической обработки семена промывали водой шесть раз по 30 мин. Обработанные семена помещали на влажную фильтровальную бумагу и помещали при 4 ° C на 48 часов, а затем переносили в комнатную температуру и давали прорасти перед пересадкой в почву.В контроле 68% семян проросли и дали плодородные растения по сравнению с 99% прорастанием необработанных семян, что указывает на то, что погружение семян в течение нескольких часов снижает их жизнеспособность. Скрининг на мутанты проводили по мутагенезу на быстрых нейтронах.
Molecular Biology
Выделение геномной ДНК проводили с использованием метода бромида цетил-триметиламмония, как описано ранее (Long and Coupland, 1998). Секвенирование генов-кандидатов проводили на мутантных растениях и на родительских растениях дикого типа, используемых для мутагенеза. MOS1 праймеры для амплификации геномной ДНК, секвенирования и генотипирования были следующими: прямой, 5′-CCCCTTGTAGCTTAGCTCCTC-3 ‘; обратный, 5’-GGCGTAGACGAAGTTGGTG-3 ‘; вперед, 5’-GACCCGACGACCAAGGAG-3 ‘; обратный, 5’-CGTCCGACACCGAGTACC-3 ‘; вперед, 5’-AATCATCACACTGCCTGCAT-3 ‘; вперед, 5′-CGAGGACTTCTTCTGCTTCTC; и обратный, 5’-TGCATGCATATGGAAACGTAG-3 ‘. MOS1 (Bradi1g18580) праймерами, использованными для количественной ОТ-ПЦР, были 5′-CGTACGGGTACGGCATGAT-3 ‘и 5′-CAGTGGGAGAGGAAGCTGAA-3’. Праймерами BdACTIN2 (Bradi1g10630), использованными для количественной ОТ-ПЦР, были 5′-GTCGTTGCTCCTCCTGAAAG-3 ‘и 5′-ATCCACATCTGCTGGAAGGT-3’.Косегрегация полиморфизма вставки в растениях Bradi1g18580 и фенотипических мутантных растениях mos1 была определена путем анализа потомства дикого типа и мутантного потомства из семи независимых линий M2. Количественный анализ ОТ-ПЦР проводили с использованием девяти меристем соцветий, срезанных с 18-дневных растений. Тотальную РНК экстрагировали из ткани соцветия с использованием реагента TRI (Sigma-Aldrich) и ДНКазу, обработанную и очищенную на колонке (Qiagen) перед синтезом комплементарной ДНК с использованием обратной транскриптазы SuperScript III (Invitrogen).Количественную ОТ-ПЦР проводили с использованием набора для количественной ОТ-ПЦР SYBR Green JumpStart (Sigma-Aldrich). Количественные реакции RT-PCR проводили в трех повторностях. BdACTIN2 (Bradi1g10630) использовали для нормализации экспрессии гена.
Гистология и гибридизация in situ
Для анализа стадий развития соцветий верхушки растений фиксировали в 4% (мас. / Об.) Параформальдегиде, дегидратировали в этаноле и заливали воском. Вложенный материал был разрезан, очищен и окрашен 0.1% (мас. / Об.) Толуидиновый синий. Гибридизацию РНК in situ проводили, как описано ранее (Long et al., 1996). Для BdKN1 ген-специфический фрагмент длиной 747 п.н. был амплифицирован из комплементарной ДНК с праймерами 5′-CATCTCCCACCCCATTTACCCT-3 ‘и 5′-TAGAGCCCACCATCGTTGATGA-3’ и клонирован в вектор pCR-Blunt II-TOPO (Invitrogen) . Антисмысловой зонд получали из этого клона транскрипцией in vitro фрагмента BdKN1 в присутствии дигоксигенин-UTP и последующим гидролизом на более мелкие фрагменты длиной приблизительно 150 п.н. перед использованием в гибридизации.
SEM
Чтобы подготовить ткань для SEM, с растений вырезали верхушки и удаляли окружающую ткань листьев, чтобы обнажить меристемы. Вершины устанавливали в горизонтальном положении на алюминиевые стержни и погружали в жидкий азот. После замораживания образцы сразу загружали в криокамеру криопереносной системы ALTO 2500 (Gatan), присоединенной к сканирующему электронному микроскопу Zeiss Supra 55 VP FEG. Образцы напыляли платиной и отображали при 3 кВ.
Выравнивание последовательностей и филогенетический анализ
Аминокислотные последовательности KNOX и BD1 были проанализированы с использованием MEGA версии 5. Последовательности были выровнены с использованием ClustalW, и филогенетическое дерево было построено с использованием алгоритма максимального правдоподобия с моделью эволюции белка JTT + G и 1000 реплик начальной загрузки. Эта модель была определена как наиболее подходящая с помощью Protest 3 (Darriba et al., 2011). Было показано, что гены KNOX класса I растений являются монофилетическими (Kerstetter et al., 1994; Malcomber et al., 2006), поэтому дерево было укоренено с помощью белка KNOX Arabidopsis ( Arabidopsis thaliana ) класса II Arabidopsis , связанного узлом -like7 (KNAT7) в качестве внешней группы. Дерево BD1 было укоренено родственным белком PUCHI (AT5G18560) из Arabidopsis и включало BD1 и близкородственный ген кукурузы ( Zea mays ) ZM2G458437 (Chuck et al., 2002), FZP, Sb02g042400, предсказанный белок в ячмень ( Hordeum vulgare ) morex-contig-1558332; и Bradi1g18580 (MOS1).Номера доступа GenBank для Arabidopsis, кукурузы и риса ( Oryza sativa ) последовательности белка KNOX были следующими: BREVIPEDICELLUS, AEE82597.1; GNARLEY1, AAP76320.1; KN1, AAP21616.1; KNAT2, AEE35073.1; KNAT6, AEE30380.1; KNAT7 AF308451; KNOX3, ACG37067.1; KNOX8, ACF81686.1; KNOX10, ACF85627.1; LIGULELESS4a, AAP31409.1; LG3, AAD13611.1; ОШ2, БАА03959.1; ОШ4, БАА79223.1; ОШ6, BAA79224.1; ОШ20, ААР87192.1; ОШ25, БАА31688.1; ОШ53, БАА79225.1; OSH71, BAA79226.1; ШЕРОХОВАЯ ОБОЛОЧКАh2, AAA86287.1; и СТРЕЛЯТЬ БЕЗУПРЕЧНЫМ, AEE33958.1. Номера доступа в GenBank для последовательностей белков BD1 / FZP кукурузы и риса были следующими: BD1, ACG41675.1; ZM2G458437, DAA41703.1; и FZP, BAC79264.1. Последовательности Brachypodium, Bradi1g07247, Bradi1g10047 ( BdKN1 ), Bradi1g12677, Bradi1g12690, Bradi1g57607, Bradi2g11540, Bradi2g38390 и Bradi1g18580 (MOS1) были идентифицированы из сборки последовательностей Vogel. Последовательности BD1 сорго ( Sorghum bicolor ) и ячменя были идентифицированы из геномных сборок (Paterson et al., 2009; Mayer et al., 2012).
Дополнительные данные
Следующие материалы доступны в онлайн-версии этой статьи.
Благодарности
Мы благодарим Эрика Фоллбрехта и Зака Липпмана за комментарии к рукописи; Саймону Хо за помощь в филогенетическом анализе; Дэвид Гарвин за предоставленные семена Brachypodium Bd21; Jozsef Palfalvi для обработки семян быстрыми нейтронами; Саре Коллиер за техническую помощь; и Эндрю Дэвис за фотографию растений.
Сноски
Автор несет ответственность за распространение материалов, составляющих выводы, представленные в этой статье, в соответствии с политикой, описанной в Инструкциях для авторов (www.plantphysiol.org): Мэри Э. Бирн (mary.byrne {at} sydney.edu.au).
www.plantphysiol.org/cgi/doi/10.1104/pp.112.212340
↵1 Эта работа была поддержана Сиднейским университетом, Исследовательским советом биотехнологии и биологических наук и Королевским обществом.
↵ [C] Некоторые рисунки в этой статье отображаются в Интернете в цвете, а в печатном издании — в черно-белом.
↵ [OA] Статьи в открытом доступе можно просматривать в Интернете без подписки.
↵ [W] Онлайн-версия этой статьи содержит данные только для Интернета.
Глоссарий
- SEM
- сканирующая электронная микроскопия
- EMS
- этилметансульфонат
- Gy
- Серый
- RT
- фактор обратной транскрипции ER
- 907 das 907 das 907 das
- Получено 5 декабря 2012 г.
- Принято 24 января 2013 г.
- Опубликовано 25 января 2013 г.
Травяные конструкции | Информационная система по кормам
Номенклатура растительных и цветочных структур
Знание названий конкретных частей растений обеспечит основу для углубленного обсуждения механизмов, которые отвечают за рост и восстановление после дефолиации. Схема и определения разделены на три категории: вегетативные, цветочные (репродуктивные) и воротничковые. Во время раннего роста, когда травянистое растение является вегетативным, идентификация затруднена.Когда видны цветочные структуры, идентификация легче. Область воротника полезна для идентификации и содержит полосу вставочной меристемы, которая при надлежащей защите будет учитывать повторный рост после скашивания или выпаса.
Вегетативные структурыОбратите внимание на следующие обозначенные элементы на чертеже:
- цветонос: самый верхний сегмент стебля, поддерживающий семенную головку.
- флаговый лист: самый верхний лист стебля, закрывающий семенную головку на стадии загрузки.
- Узел стебля : сплошная область на стебле, которая дает начало листовой оболочке. У некоторых видов трав в нижних узлах стебля могут появиться придаточные почки, способные давать новые побеги.
- стебель: центральная ось побега зрелой травы, состоящая из узлов и междоузлий; каждый узел несет лист.
- пластинка листа: часть листа над оболочкой, также известная как пластинка.
- влагалище листа: нижняя часть травы, окружающая ее междоузлия в стебле.
- ушные раковины: короткие, часто когтеобразные придатки у основания листовой пластинки, которые стремятся обхватить влагалище в междоузлиях стебля. Различные формы могут быть полезны для идентификации определенных трав.
- язычок: различно модифицированное продолжение ножен, лежащее у основания лезвия; часто это вертикальная перепонка, а в некоторых случаях — просто щетина.
- крона: базальная зона побега, источником которой была ткань у основания колеоптиля на стадии проростка.Крона важна для многолетнего роста растения, так как эта зона состоит из зимующих тканей (базальных междоузлий, корневищ, столонов, клубнелуковиц), которые дают новые побеги следующей весной. У однолетних трав крону не развивают.
- столон: распростертый или ползучий надземный стебель, укореняющийся в узлах; средство вегетативного размножения.
- корневище: распростертый подземный стебель, способный укореняться в узлах и становиться прямостоячим на вершине; средство вегетативного размножения.
- Культиватор : дочернее растение, побег, способный дать новое растение.
- Меристема : клеток, способных к росту.
- воротниковая область: Воротниковая область листа является наиболее полезной областью для идентификации трав вегетативной стадии. Позже, после появления всходов семян, цветочные структуры могут стать более очевидным средством идентификации. Воротничковая область состоит из листовой пластинки, оболочки листа, обертывающей стебель, ушных раковин (если они есть), язычка и соединительной ткани, называемой воротничком.Каждый вид уникален наличием, размером и формой ушек и язычков в этой листовой зоне. Воротник (соединительная ткань) представляет собой узкую полосу вставочной меристемы (ткани, способной к росту), которая отвечает за рост лопатки. У незрелых лопастей эта меристема обеспечивает дальнейший рост лопастей после дефолиации.
На первых фотографиях виден воротник с пластинкой (пластинка), ушные раковины, язычок и влагалище листа.
- полотно (пластинка): часть полотна над воротником
- воротник: тонкая полоса меристематической ткани на стыке листовой пластинки (пластинки) и влагалища
- ушных раковин: когтеобразных придатков у основания лезвия некоторых трав (некоторые травы не имеют ушных раковин)
- язычок: вырост на внутреннем стыке влагалища листа и пластинки, часто перепончатый, иногда бахрома из волосков
- оболочка: нижняя часть створки, закрывающая междоузлия
На средней фотографии показан воротничок, на котором видна вставочная меристематическая ткань (более светлая) прямо под верхней пластинкой листа.Нижняя пластинка листа загнута назад, показывая язычок, но на краю складки видна часть беловатого воротничка.
На нижнем снимке — воротничок — узкая полоса меристематической ткани тимофеевки.
Область воротника также важна при выращивании травы. При надлежащей защите эта полоса вставочной меристемы будет обеспечивать непрерывный рост лезвий после скашивания или выпаса незрелых листовых пластин.
Цветочные структуры- соцветие: цветочная головка, оканчивающаяся стеблем, состоящая из набора цветков, расположенных на общей оси. Существует три основных типа соцветий травы: 1. метелка, 2. колос, 3. кисть.
- rachis: центральная ось высевающей головки.
- колоск: цветковая единица, состоящая из одного или нескольких цветков, окруженных двумя чешуйками (прицветниками). Когда колоски прикреплены непосредственно к позвоночнику, соцветие называется колосом (пшеница, рожь, ячмень, райграс).Когда колоски прикрепляются к рахису короткими цветоножками, соцветие называется кистью. Когда колоски прикрепляются с помощью веточки, соцветие называется метелкой.
- Цветоножка : у злаков, короткий сегмент стебля, поддерживающий колоск. Такие колоски называют педицеллятными.
- чешуек: прицветников, окружающих цветочек (и). Колоск можно охарактеризовать как «пару» чешуек с закрытым цветком (цветами). Наружная (нижняя) чешуйка всегда самая большая из пары.У некоторых видов (семейство Paniceae) самая верхняя чешуйка сильно сокращена и в значительной степени заменена леммой о стерильном цветке, содержащемся внутри.
- цветочек: мелкий цветок; репродуктивная единица колоска травы, состоящая из леммы и палеи, и небольшого цветка, который они содержат (см. колоск).
- rachilla: сегментированная центральная ось колоска выступает в колосках, несущих два или более плодовитых цветков. Каждый сегмент рахиллы несет соцветие, поэтому в обмолоченной форме одно семя обычно сохраняет сегмент или сустав рахиллы.Наличие или отсутствие сегмента рахиллы позволяет распознать многие семена. Кроме того, форма и размер сегмента сильно различаются у разных видов.
- lemma: — более крупный внешний прицветник, который вместе с палеей служит для удерживания цветков, удерживаемых внутри. Lemma и palea обеспечивают защитное покрытие для развивающихся цветков, а также для семян после созревания.
- ость: волокнистая щетина (часто называемая бородой), которая является продолжением средней жилки леммы.Он может возникать на кончике леммы или на абаксиальной (внешней) поверхности под кончиком.
- борода: общий термин для ости.
- palea: более короткий верхний прицветник, который, наряду с леммой, служит для удерживания цветков, удерживаемых внутри (см. Лемму).
- тычинок: мужской орган цветка, поддерживающий пыльники, производящие пыльцу.
- пыльник: часть тычинки, несущая пыльцу, состоящая из одного или двух пыльцевых мешочков.
- пыльца: структуры, возникающие в результате созревания микроспоры.
- пестик: женский орган цветка, состоящий из рыльца, стебля и завязи. На рыльце образуется пыльцевое зерно, которое при прорастании дает пыльцевую трубку, которая проходит через рыльце в завязь.
Неродные крабовые травы (род Digitaria) Нью-Йорка
В штате Нью-Йорк произрастают три неместные крабовые травы: Digitaria ciliaris, D.ischaemum, и D. sanguin alis . Эти однолетние травы начинают цвести в середине лета и продолжают цвести и плодоносить, пока мороз не отбросит их назад. Родной ареал D. ciliaris неясен. Недавние авторы (например, Wipff 2003, Shouliang and Phillips 2006) считали, что он широко распространен в тропических и умеренно-тропических частях мира. Digitaria ischaemum и D. sanguinalis произрастают в Евразии (Wipff 2003). Все три вида широко распространены по всему Нью-Йорку и встречаются в различных нарушенных естественных и чужеродных средах обитания.Все они внешне похожи тем, что имеют длинные пальцеобразные ветви соцветий, расходящиеся от вершины или около вершины стебля.
Соцветие Digitaria ciliaris
Каждая ветвь соцветия имеет многочисленные колоски на коротких цветоножках.
Ветка соцветия Digitaria ischaemum
При более внимательном рассмотрении оказывается, что колоски сгруппированы вместе.У D. ischaemum колоски сгруппированы по три, а у D. ciliaris и D. sanguinalis они сгруппированы попарно. Парные колоски двух последних видов имеют колоски на цветоножках разной длины, один очень короткий, а другой более длинный.
Digitaria ischaemum с колосками, расположенными группами по три. Некоторые колоски были удалены, а некоторые разложены друг от друга для удобства наблюдения.
Digitaria ciliaris с парными колосками. Некоторые колоски были разнесены друг от друга для удобства наблюдения.
Digitaria ischaemum также сильно отличается от двух последних видов во многих других отношениях, включая:
Digitaria ischaemum | Digitaria ciliaris и D.sanguinalis | |
2-я лемма | черный, темно-фиолетовый или темно-коричневый, в основном скрытый за второй чешуей | желтый, зелено-серый или серо-голубой, часто с пурпурным оттенком, верхняя часть хорошо видна |
2-я чешуя | длиной или немного короче второй леммы | примерно на 40-75% до 2-й леммы |
1-я чешуя | отсутствует или рудиментарен, если присутствует тонкая полупрозрачная усеченная мембрана ≤ 0.Длиной 2 мм; | маленькие, но видимые, толстые, непрозрачные, длиной 0,1-0,8 мм |
колоски | длина 1,8-2,3 мм | длина 2,3-4,0 мм |
Колоск Digitaria ischaemum
Колоск Digitaria ciliaris
Как только вы познакомитесь с этими видами, вы увидите относительно небольшие колоски D.ischaemum позволяет отличить его от двух других видов на расстоянии. При необходимости быстрое более тщательное изучение растений может подтвердить определение.
Digitaria ciliaris и D. sanguinalis — очень похожие виды, и их легко спутать. Лучше всего их различить, наблюдая за боковыми жилками первых лемм. У D. ciliaris боковые жилки гладкие или иногда с несколькими небольшими спикулами, а у D.sanguinalis многие боковые жилки имеют шероховатость, по крайней мере, в верхней половине. Шероховатую текстуру, образованную множеством мелких спикул, трудно различить без хорошего освещения и увеличения.
Колоски Digitaria sanguinalis (слева) и D. ciliaris (справа)
Кроме того, верхняя поверхность самых верхних листовых пластинок D. ciliaris иногда бывает шероховатой, но в остальном голая или имеет только несколько волосков на основе папиллозы, особенно у основания по сравнению с верхней поверхностью самой верхней листовой пластинки, умеренно или густо опушенной с волоски на основе папиллозы в D.sanguinalis .
Верхняя поверхность пластинок листьев Digitaria sanguinalis (вверху) и D. ciliaris (внизу)
Есть и другие отличия, такие как относительная длина второй чешуйки по отношению к колоску и абсолютная длина колоска, хотя есть некоторые совпадения с этими признаками. А в Нью-Йорке я иногда встречал промежуточные звенья. Сообщалось о промежуточных растениях из других областей, и предполагалось, что они являются гибридами или представляют некоторый уровень интрогрессии между двумя видами (Ebinger 1962, Gould 1963, Webster and Hatch 1981).
Первая лемма из Digitaria ciliaris и D. sanguinalis обладает интересными волосками. Основные волосы мягкие, гибкие и белые (иногда переходящие в бледно-белые или бледно-коричневые). В молодом возрасте эти волоски плотно сбиты вместе и прижаты к поверхности леммы. В основном они встречаются в двух местах, по краям и между двумя внутренними боковыми жилками. В молодом возрасте они лежат параллельно жилкам и выглядят как другая жилка, но в зрелом возрасте они вздуваются и выступают за поверхность чешуи, их кончики часто соединяются.
Колоски Digitaria ciliaris незрелые (слева) и зрелые (справа)
Помимо мягких белых волосков, первые леммы Digitaria ciliaris и D. sanguinalis иногда также имеют жесткие стекловидные желтые волоски, хотя я не видел экземпляров D. sanguinalis из Нью-Йорка с такими волосы. Когда они присутствуют, жесткие стекловидные желтые волосы обычно встречаются с гибкими белыми волосками между двумя внутренними боковыми жилками и обычно выступают далеко за пределы гибких белых волосков.
Колоск Digitaria ciliaris
В ботаническом сообществе нет единого мнения относительно того, достойны ли растения D. ciliaris с жесткими стекловидными желтыми волосками таксономического признания. Хенрард (1950) в своей всемирной монографии по роду Digitaria определил эти растения в конкретном ранге как D. chrysoblephara . Wipff (2003), рассматривая Digitaria во флоре Северной Америки к северу от Мексики, распознал эти растения как D.ciliaris var. chrysoblephara , но отметил, что необходимы дальнейшие исследования. Shouliang и Phillips (2006) во Флоре Китая отметили, что эту «форму» иногда выделяли как разновидность. Вильхальм (2009) в своем обзоре D. ciliaris в Европе считал, что растения с жесткими стекловидными желтыми волосками представляют только форму. Велдкамп (1973) в своей редакции Digitaria Malesia считал, что D. ciliaris никогда не обладает желтыми стекловидными волосками, и считал такие растения частью изменчивости близкородственных D.bicornis. Webster (1987) в своем пересмотре видов, тесно связанных с D. ciliaris из Северной Америки, признал, что D. ciliaris могут быть вариабельными в зависимости от наличия или отсутствия стекловидных желтых волосков, но не дали таксономического признания растениям. из D. ciliaris с такими волосками. Что касается применения названий, Вебстер рассмотрел название D. ciliaris var. chrysoblephara является синонимом D. bicornis , в то время как Wipff (2003) и Shouliang and Phillips (2006) относят его к D.ciliaris .
У экземпляров D. ciliaris из Нью-Йорка, которые я наблюдал, когда они присутствовали, обычно не так много этих стекловидных желтых волосков на каждую лемму, и иногда кажется, что у многих лемм эти волоски отсутствуют, в то время как у других есть только один или два. Трудно точно оценить, есть ли у особи такие волосы, потому что эти волосы приобретают желтый цвет по мере созревания, а когда незрелые, смешиваются с белыми гибкими волосками и прижимаются ими к телу чешуи, что затрудняет их различение.В 2020 году я собрал десять экземпляров D. ciliaris из западных, центральных, северо-западных и северо-восточных регионов Нью-Йорка. При тщательном изучении этих экземпляров выяснилось, что все они имеют по крайней мере несколько жестких стекловидных желтых волосков. Похоже, это противоречит утверждению Випффа (2003) о том, что растения D. ciliaris без жестких стекловидных желтых волосков более распространены в Северной Америке к северу от Мексики, чем растения с такими волосками.
D igitaria ischaemum и D.sanguinalis давно известны как обычное явление в Нью-Йорке (например, Хаус [1924]). С другой стороны, Смит (1965) в своем Контрольном списке трав Нью-Йорка, возможно, был первым, кто сообщил о D. ciliaris (как D. ascendens ) из Нью-Йорка. Он отметил, что его нашли только в качестве беспризорника в юго-восточной части штата. Карточки Музея штата Нью-Йорк, серия больших манильских карточек, которые ботаники Музея Нью-Йорка использовали для записи и картирования распространения видов в штате, отражают оценку Смитом вида в то время; карта музейной карты штата Нью-Йорк для номера D.ciliaris показаны три местонахождения этого вида, все в юго-восточной части штата.
Музейная карта штата Нью-Йорк карта Digitaria ciliaris , на которой показаны три популяции (черные точки), которые ограничены юго-восточной частью штата.
Но Digitaria ciliaris с тех пор стала широко распространенной и распространенной во многих, если не во всем штате; между 2003 и 2020 годами я собрал этот вид во всех регионах штата, и его никогда не было сложно найти.
Карта Нью-Йорка, показывающая, где я собирал гербарные купоны на Digitaria ciliaris . Обратите внимание, что я наблюдал этот вид в других местах штата, и есть образцы гербария, документирующие и другие популяции.
Похоже, что этот вид быстро распространился в штате в недавнем прошлом, хотя для подтверждения этой гипотезы необходим тщательный обзор гербарных образцов. Яцкевич (1999) сообщил о подобном недавнем (с 1963 г.) распространении этого вида в штате Миссури. Digitaria ciliaris , вероятно, распространилась за пределы Нью-Йорка в некоторые регионы, которые еще не зарегистрировали этот вид, такие как Онтарио (Oldham, 2017) и Вермонт (Gilman, 2015).
В Нью-Йорке, аналогично D. ischaemum и D. sanguinalis , D. ciliaris произрастает в естественных нарушенных средах обитания, таких как гравий и песчаные отмели в реках и в зонах впадения прудов, озер и рек. а также на нарушенных человеком участках, таких как сельскохозяйственные поля, обочины дорог и трещины на тротуарах в городских районах.
Среда обитания Digitaria ciliaris в Нью-Йорке. Просадка на берегу озера Шамплейн, Clinton Co., NY
Среда обитания Digitaria ciliaris в Нью-Йорке. Осадка на краю залива Каюга, Tompkins Co., NY
Среда обитания Digitaria ciliaris в Нью-Йорке. Деревня Дружбы, Allegany Co., NY
Среда обитания Digitaria ciliaris в Нью-Йорке.Сельскохозяйственное поле в Tompkins Co., NY
Последнее замечание. При подготовке этого сообщения в блоге я понял, что некоторые из моих недавних коллекций D. ciliaris , по-видимому, имеют по крайней мере некоторые характеристики близкородственных, а также спорных видов D. bicornis. Webster and Hatch (1981) предоставили некоторые доказательства того, что эти два вида отличаются друг от друга, и многие авторы продолжают признавать их как таковые, хотя и с разными описаниями (Veldkamp 1973, Wipff 2003, Shouliang and Phillips 2006, Weakley 2015, Boonsuk et al.2016). Другие настроены более скептически (Wilhalm 2009). D igitaria bicornis , как сообщается, широко распространена в тропиках и субтропиках мира (Webster 1987), а также обычна на прибрежных равнинах на юго-востоке Соединенных Штатов, возможно, произрастая на севере до Вирджинии или Мэриленд (Wipff, 2003). Очевидно, что необходима современная ревизия комплекса D. ciliaris . Так что следите за новостями, чтобы узнать больше о крабовой траве!
Цитированная литература:
Бунсук, Б., П. Чантаранотаи и Т. Р. Ходкинсон. 2016. Таксономическая ревизия рода Digitaria (Panicoideae: Poaceae) в континентальной части Юго-Восточной Азии. Phytotaxa 246: 248–280.
Эбингер, Дж. Э. 1962. Действительность видов травы Digitaria adscendens . Бриттония 14: 248–253.
Гилман А. В. 2015. Новая флора Вермонта. Воспоминания о ботаническом саду Нью-Йорка 110: 1–615.
Gould, F. W. 1963. Цитотаксономия Digitaria sanguinalis и D.Адсенденс . Бриттония 15: 241–244.
Хенрард, Дж. Т. 1950. Монография рода Digitaria . Universitaire Pers Leiden, Лейден, Нидерланды.
House, H. D. 1924. Аннотированный список папоротников и цветковых растений штата Нью-Йорк. Бюллетень Музея штата Нью-Йорк 254. Университет штата Нью-Йорк, Олбани, штат Нью-Йорк, США.
Олдхэм, М. Дж. 2017. Список сосудистых растений Каролинской зоны Онтарио (экорегион 7E). Каролинская Канада и Министерство природных ресурсов и лесного хозяйства Онтарио, Питерборо, Онтарио, Канада.
Шоулян К. и С. М. Филлипс. 2006. Digitaria Haller. Страницы 539–547 в З. Ю. Ву, П. Х. Рэйвен и Д. Ю. Хонг, редакторы. Флора Китая. Vol. 22 (Poaceae). Пресса Ботанического сада Миссури, Сент-Луис, Миссури, США.
Смит, С. Дж. 1965. Контрольный список трав штата Нью-Йорк. Бюллетень Музея штата Нью-Йорк 403. Университет штата Нью-Йорк. Государственный департамент образования, Олбани, штат Нью-Йорк, США.
Велдкамп, Дж. Ф. 1973. Пересмотр Digitaria Haller (Gramineae) в Малезии.Заметки о малезийских злаках VI. Блумеа 21: 1–80.
Уикли А. С. 2015. Флора южных и среднеатлантических штатов. Рабочий проект от 21 мая 2015 г. Гербарий Университета Северной Каролины, Ботанический сад Северной Каролины, Университет Северной Каролины в Чапел-Хилл, Чапел-Хилл, Северная Каролина, США.
Вебстер Р. Д. 1987. Таксономия Digitaria секции Digitaria в Северной Америке (Poaceae: Paniceae). Сида 12: 209–222.
Вебстер Р. Д. и С. Л. Хэтч.1981. Таксономические отношения техасских образцов Digitaria ciliaris и Digitaria bicornis (Poaceae). Сида 8: 34–42.
Wilhalm, T. 2009. Digitaria ciliaris в Европе. Willdenowia 39: 247–259.
Wipff, J. K. 2003. Digitaria Haller. Страницы 358–383 в Флоре Северной Америки Редакционный комитет, М. Э. Баркуорт, К. М. Капелс, С. Лонг и М. Б. Пип, редакторы. Флора Северной Америки к северу от Мексики, том 25, Magnoliophyta: Commelinidae (частично): Poaceae, часть 2.Издательство Оксфордского университета, Нью-Йорк, Нью-Йорк, США.
Яцкевич Г. 1999. Флора Штайермарка в штате Миссури. Том 1. Исправленное издание. Департамент охраны окружающей среды штата Миссури, Джефферсон-Сити, Миссури, США.
ГРИН-Глобал
Национальная политика США по распространению зародышевой плазмы растенийГермоплазма растений распространяется среди ученых, преподавателей, производителей и других добросовестных исследовательских и образовательных организаций из U.S. Национальная система зародышевой плазмы растений (NPGS) активна сайты сбора. Куратор NPGS и / или руководитель исследования будет, в соответствии с действующими политиками и процедурами NPGS, при необходимости определить законность запроса.
Раздача для выполнения запросов на репатриацию подвыборок коллекций зародышевой плазмы в страну или общину происхождения, особенно после природных или техногенных катастроф, считаются приоритетными.
Хотя распределения для исследований, образования и репатриации являются наивысшим приоритетом, NPGS также поощряет различные организации по сохранению семян и общественные сады для проведения мероприятий по сохранению зародышевой плазмы, в которых участвуют многие отдельные лица и группы по всей стране. Элементы NPGS сотрудничают с организациями по сохранению семян и общественными садами и могут хранить зародышевую плазму для и распространять зародышевую плазму среди таких организаций.
Распространение гермоплазмы из коллекций NPGS для выполнения запросов от лиц, ищущих бесплатную зародышевую плазму строго для домашнего использования обычно считается ненадлежащим использованием ограниченных ресурсов и противоречит политике правительства США не конкурировать с коммерческие предприятия. Подателей запросов можно соответствующим образом попросить обосновать использование конкретной зародышевой плазмы NPGS вместо этого. подходящей коммерчески доступной зародышевой плазмы.
Образцы, перечисленные в базе данных информационной сети о ресурсах зародышевой плазмы (GRIN) как «недоступные» из-за недостаточного или низкого семена жизнеспособности и / или запланированные к регенерации, как правило, не будут доступны для распространения.
Другие образцы перечислены в GRIN как «недоступные», потому что они не являются частью коллекции NPGS как таковой, но сохраняются в NPGS. генные банки для удовлетворения конкретных потребностей, как описано ниже в разделе «Категории распространяемой и доступной зародышевой плазмы».” В эту категорию входят определенные образцы улучшенной зародышевой плазмы, которые доступны только у владельца / разработчика. Другие присоединения требуют, чтобы запрашивающая сторона выполняла определенные условия, прежде чем распространение станет возможным.
Сайты NPGS не будут распространять зародышевую плазму на международном уровне, если они не могут соответствовать требованиям ввоза или карантина. страны-получателя, если запрашивающая сторона не может предоставить действительный отказ от таких требований.
Способы вязания узора «Азиатский колосок»
.Содержимое
Удивительно интересный рисунок азиатских колосков, которые можно соединить с помощью самых простых видов петель. Вещь получается объемной и с первого взгляда привлекает к себе внимание.
Простой способ сделать азиатский колосок
Узор можно выполнить двумя способами.По первому из них мы свяжем ряды по частям, то есть закроем часть петель, а затем снова наберем. По второму — в одном ряду необходимо выполнить набор и закрытие петель, а узор выполняется отдельно.
Количество провязанных рядов и оригинал Количество петель на спицах может быть совершенно разным. Стоит помнить, что в готовом виде размер изделия будет в 1,5 раза меньше, поэтому его необходимо учитывать при расчете длины и ширины ткани.
Азиатский узор №1
Начнем с 40р. 1-6П. Вяжем лицевой гладью 6р. Если хотите получить более нежный узор, можно сделать колоски менее тонкими, связав 3-5р лиц. Гладкий.
Напомним, что при выполнении этого простого шаблона необходимо связать нечетные ряды ЛП, а четные — ПЭ.
7р .: Находим центральные узоры вязания и закрываем их. То есть выполняем 10LP. 11-я точка пересчитывается на правую спицу, мы помещаем после нее рабочую нить, затем перемещаем 12-ю точку и закрываем с 12 по 11.По этой схеме закрываем все 20п. Далее — 10LP.
8Р .: 10ИП, 20п — набираем, 10ИП. После него общее количество петель на спицах 40.
Повторяем 1-8П. Эта схема подразумевает любое количество повторений раппорта. В результате у вас должно получиться полотно, как показано на картинке. Выполнить его несложно, мастер подключит за 20-30 минут.
Как набирать петли Есть несколько способов набрать на спицы. Азиатский узор будет выглядеть аккуратнее, если использовать следующую технику:
- Две последние петли левой спицы раздвинуты;
- Между ними протягиваем нить;
- Надеваем на левую спицу.Получили первую петлю;
- Осталось повторить этот процесс еще 19 раз и все 20р будут набраны;
- Чтобы связать их с основным вязанием, нужно сделать 21н и вернуть на правую спицу.
Такой прием позволяет получить более плотное полотно, которое станет отличной основой для формирования колосков.
Процесс сборки колосов
Теперь колоски нужно собрать в единое целое. Для этого переверните изделие изнаночной стороной.
Первый колосок получается скручиванием первой и второй полос. Далее третью полоску протягивают через петлю второй полоски и так до конца полотна.
Чтобы колосок получился красивее и нежнее, нужно максимально растянуть каждую полоску и дырочку. Переверните вязание на лицо и расправьте каждый колоск, сформируйте его. Видео этого процесса можно посмотреть на нашем сайте.
Азиатский узор №2
Для начала во втором режиме вязания определяем ширину колоска.Наберем именно такое количество петель. Если колоск 6н, то начинаем с этого количества. Хотя это может быть 10n, 8n, 4n, в зависимости от того, какие узоры вы хотите получить. Просто класс, как выглядят большие лопатки на шарфах и палантинах, для блузок лучше набрать 4-6п. Немаловажную роль играет толщина пряжи.
Мы помним, набирая спицами, что на этом рисунке не учитываются края.
Видео: Уроки вязания для начинающих
Колоск спереди
Круглый циферблат 18р., А потом по этой схеме:
- IP: только LP;
- 2П .: только ИП. Это основа будущего.
Схема следующая:
- 3р .: начинаем вязать первое полотно. Его размеры: ширина — 6п, высота — 10п .;
- 5ЛП, перевернуть изделие, 6ИИП. По этой схеме вяжем еще 8 рядов. Первое лезвие готово.
Для перехода к следующему элементу необходимо (по счету будет 13р) соединить 6ЛП с правой спицы и 3п с левой.Изделие перевернуто. 14Р .: 6ИП, 3п — не связывать. Дальше по схеме 3-13П только 6н. Третий элемент должен начинаться с 23P.
Данную схему проводим до тех пор, пока не будет достигнута необходимая длина изделия. В итоге у вас должно получиться 6п.
Теперь, прежде чем перейти к другой стороне колоска, нужно выполнить промежуточные ряды. Сначала будет серия IP, затем LP. Вы можете повторить их снова. Но нужно закончить это рядом LP.
Колоск с изнанки
Изделие переворачиваем и начинаем вязать изнаночную сторону.Просто класс, если делать все с первого раза, так как узор довольно сложный. Теперь нам нужно направить колоск в другую сторону, чтобы узор получился симметричным и красивым. Мастер сразу поймет, что схема здесь будет аналогична предыдущей стороне.
Запускаем первый элемент:
- 1Р .: 1Кр, 5ИП, полотно переворачиваем, 6ЛП .;
- 2-7р .: продолжаем по этой технологии. Получили первый элемент;
- СП: вяжем 9н (6н + 3н от основного полотна), все ИП;
- 9Р.: переворачиваем полотно. Снова выполняем чередование IP и LP.
В последнем ряду соединяем все петли и закрываем их. При правильном приклеивании колоски будут смотреть в разные стороны от центра. Если такой рисунок у вас получился, то получился азиатский узор.
Видео: Вязание азиатского колоска без дырочки